
Abstract
Cord blood-derived endothelial colony-forming cells (CB-ECFCs) hold significant promise for regenerative medicine due to their unique vasculogenic and immunomodulatory properties. These cells exhibit a superior proliferative capacity, robust ability to form vascular networks, and lower immunogenicity compared to adult and embryonic stem cell-derived counterparts. The immune-privileged characteristics of CB-ECFCs, including reduced expression of pro-inflammatory mediators and tolerance-inducing molecules such as HLA-G, further enhance their therapeutic potential. Their low immunogenicity minimizes the risk of immune rejection, making them suitable for allogenic cell therapies. Their application extends to complex tissue engineering and organ revascularization, where their ability to integrate into three-dimensional scaffolds and support vascular tree formation represents a significant advancement. Moreover, CB-ECFCs’ capability to adapt to inflammatory stimuli and retain immunological memory highlights their functional versatility in dynamic microenvironments. This review highlights the remarkable ontogeny of ECFCs while unveiling the unparalleled potential of CB-ECFCs in revolutionizing regenerative medicine. From pre-vascularizing engineered tissues and organoids to pioneering cell-based therapies for cardiovascular, dermatological, and degenerative diseases, CB-ECFCs stand at the forefront of cutting-edge biomedical advancements, offering unprecedented opportunities for therapeutic innovation. By leveraging their vasculogenic, immune-regulatory, and regenerative capacities, CB-ECFCs offer a robust alternative for addressing the challenges of vascular repair and organ engineering. Future research should focus on unraveling their transcriptomic and functional profiles to optimize clinical applications and advance the field of regenerative medicine.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The field of regenerative medicine has made significant strides in recent years, with the potential of allogenic cell therapy emerging as a promising avenue for treating a variety of conditions [1]. Among the various cell types under investigation, endothelial colony-forming cells (ECFCs) derived from cord blood have garnered particular interest due to their unique vasculogenic properties and potential therapeutic applications [2]. These cells are notable for their ability to form blood vessels and their role in vascular repair, which positions them as a critical component in developing effective cell-based therapies. After years of inconsistency, consensus has been recently obtained about their isolation and characterization from cord and adult blood [2,3,4].
Cord blood, a rich source of hematopoietic stem cells, also provides a reservoir of ECFCs with distinct advantages over other sources [5]. These cells are less mature, exhibit higher proliferative capacity and clonogenic properties, and demonstrate a robust ability to form vascular networks. Moreover, the use of cord blood-derived ECFCs (CB-ECFCs) introduces fewer ethical concerns compared to embryonic stem cells (ESCs). It also presents a lower risk of graft-versus-host disease compared to adult stem cells, highlighting their potential for safer and more effective clinical applications. A pivotal aspect of using ECFCs in allogenic cell therapy could be their immune privilege. The concept of immune privilege refers to the ability of certain cells or tissues to evade detection and destruction by the host immune system. This property is particularly beneficial for allogenic transplants, as it can reduce the likelihood of immune rejection and improve the overall success rates of cell-based therapies. Understanding and harnessing the mechanisms of immune privilege in CB-ECFCs could pave the way for more widespread and successful implementation of these cells in clinical settings.
This paper aims to explore the characteristics and therapeutic potential of CB-ECFCs, with a focus on ontogeny, immune-privileged status and bioengineering applications. We will review current research on the mechanisms that confer an immune privilege to these cells, discuss their advantages over other cell types, and evaluate the feasibility of their use in allogenic cell therapy. By delving into these aspects, we hope to provide a comprehensive overview of this field and outline the potential pathways toward translating this promising research into clinical practice.
ECFCs: a human vasculogenic cell type usable for vessel reconstruction
ECFC ontogeny
The identification of adult endothelial progenitor cells (EPCs) in humans in 1997 dramatically changed the understanding of postnatal blood vessel formation [6]. Within the different EPC subtypes, ECFCs also named as non-hematopoietic EPCs [7] have gained recognition for their notable ability to promote vasculogenesis and their culture and isolation have been standardized recently [3]. Despite the substantial interest in their application as a cell therapy product or as a component of liquid biopsies, the exact tissue and molecular origins of ECFCs remain contentious. While Lin et al., using a sex-mismatch human bone marrow transplantation model, reported that ECFCs do originate from bone marrow [8], Fujisawa et al. demonstrated the opposite more recently in five male recipients who had received allogenic bone marrow transplants from female donors to treat hematological disorders [9].
ECFCs were initially identified in peripheral blood and cord blood-derived mononuclear cells cultures [5, 8, 10]. Subsequent studies demonstrated that ECFCs can arise from CD34 + cells, as well as from specific subpopulations expressing CD146+/CD34+/CD45+/CD133 + or CD177 + markers [11,12,13]. Subsequent research conducted by Case et al. and Timmermans et al. demonstrated that although CD34+/CD45− cells possess the ability to differentiate into ECFCs, they lack the capacity to generate hematopoietic colonies [14, 15]. This was later confirmed by Tura et al., who identified CD34+/CD146+/CD133 − cells as the precursor population for ECFCs [16]. However, the exact stem cell at the origin of ECFCs is quite elusive. thus, several descriptions of ECFCs Originating from CD34 + are convincing at this time. Lin et al. [17] provides crucial insights into the developmental origins of ECFCs. Their study demonstrates that ECFCs originate specifically from CD34 + cells in umbilical cord blood and adult peripheral blood. CD34bright mononuclear cells in cord blood are the primary source of ECFCs, confirming their distinct endothelial lineage. Lineage tracing experiments in mice reveal that ECFCs are not derived from the bone marrow hematopoietic system but rather from vascular endothelium. Unlike hematopoietic EPCs, which play only proangiogenic roles, ECFCs possess real vasculogenic capacity, forming functional blood vessels in vivo [10]. The study demonstrates that ECFCs are a distinct subset of postnatal, tissue-derived vascular progenitors. Another subset of CD34 + cells has been described to give rise to ECFCs like cells. Indeed, very small embryonic-like stem cells (VSELs), identified in both humans and mice, have been proposed to give rise to differentiate into multiple lineages [18] but in particular in endothelial cells in vivo [19], in vitro [20] but also in ECFCs-like cells after differentiation in vitro [21]. VSELs have been first described as CD133+ small dormant stem cells and are linked to migratory primordial germ cells. They have been found in bone marrow and other organs, such as the heart and lungs [18]. The pluripotent capacity of VSELs presents a promising avenue for resolving the debate about bone marrow stem cell reservoirs and the conflicting data surrounding endothelial ontogeny and the Hemangioblasts hypothesis in humans. By bridging the gap between the developmental plasticity of stem cells and the observed functional properties of ECFCs, VSELs could unify disparate findings, providing a comprehensive framework to understand the ontogeny and potential reservoirs of endothelial progenitor cells. This resolution could advance both theoretical knowledge and practical applications in regenerative medicine.
Despite their potential in clinic, traditional methods of isolating and expanding CD34 + cells have been limited by scalability and reproducibility. To overcome this, Pr Hénon’s team developed an automated GMP-compliant expansion platform designed for large-scale production of expanded CD34 + cells [22] named ProtheraCytes, ensuring a controlled and standardized therapeutic product. The ProtheraCytes manufacturing process involves several key steps. CD34 + cell mobilization and collection is performed using granulocyte-colony stimulating factor to mobilize CD34 + cells from the bone marrow into the peripheral blood, followed by collection through leukapheresis. The cells then undergo a nine-day expansion process within a closed, aseptic bioreactor system known as StemXpand, increasing CD34 + cell numbers by approximately sixteen- to nineteen-fold. The final product is subjected to immunoselection and quality control, ensuring high viability, purity, and paracrine secretion potential while complying with Good Manufacturing Practice standards [22]. ProtheraCytes exhibit a gene expression profile indicative of early endothelial differentiation potential [23] https://doi.org/10.1038/s41598-023-47079-8. Transcriptomic analysis reveals upregulation of genes associated with mesoderm and endothelial commitment [23]. These expanded cells also contain an enriched content of CD34 + VSELs, which retain their pluripotency and probably contribute to endothelial differentiation. Thus, protheraCytes represent a new class of expanded CD34 + progenitor cells, designed to harness the regenerative potential of endothelial lineage cells and confirm that CD34 + cells have potential for endothelial lineage https://doi.org/10.1038/s41598-023-47079-8 [24].
Moreover, tissue-Resident Endothelial Stem Cells expressing CD157 (endothelial protein C receptor; EPCR) have been described to be a good candidate for ECFCs. Indeed, two significant papers by Yoder’s group [25, 26] provide compelling evidence that ECFCs originate from this resident endothelial populations. Wakabayashi et al. describe a population of CD157 + vascular endothelial stem cells (VESCs) found in large arteries and veins across multiple organs. Using genetic lineage tracing in mice, they demonstrated that these cells exhibit clonal proliferative capacity and are able to generate new ECs. They maintain large blood vessels and sinusoids for over a year, indicating a role in long-term vascular homeostasis. They expand and regenerate entire vasculature structures in response to injury. Furthermore, these cells can be transplanted to rescue bleeding phenotypes, confirming their regenerative capabilities. The discovery of CD157 as a marker for VESCs is particularly important, as it provides a means to prospectively isolate and study these cells, offering strong in vivo evidence that the vascular system contains its own stem cell reservoir, rather than depending solely on circulating progenitors or embryonic-like progenitors. Building on this work, Naito et al. [26] established a robust protocol for isolating tissue-resident VESCs from the mouse liver. Their findings confirm that CD157 + VESCs exist within liver blood vessels and can be efficiently isolated using fluorescence-activated cell sorting (FACS). These cells exhibit self-renewal properties and contribute to new blood vessel formation in an experimental setting. Their angiogenic potential is greater than that of mature endothelial cells, suggesting that vascular homeostasis relies on a subpopulation of stem-like cells within vessels. These two studies strongly indicate that ECFCs are likely derived from resident vascular progenitors rather than a single embryonic progenitor pool. Adult vasculature harbors its own regenerative endothelial cell population, reducing reliance on circulating or bone marrow-derived progenitors. CD157 + VESCs may serve as a therapeutic target for vascular repair. Bhagwani et al. [27] demonstrate that primitive endothelial cells can undergo clonal selection and contribute to vascular disease, particularly in pulmonary hypertension (PH). Their study found that CD117 + primitive endothelial cells proliferate abnormally, leading to occlusive vascular pathology. These cells retain endothelial characteristics but also show mesenchymal differentiation potential. This suggests that certain endothelial clones can drive pathological vascular remodeling. Lin et al. [28] identified ABCG2-expressing clonal endothelial cells as a unique population that maintains vascular homeostasis and contributes to endothelial turnover. These cells exhibit higher proliferative potential than mature endothelial cells. They are distinct from embryonic-like progenitors and instead represent a local vascular stem cell pool. Their findings reinforce the notion that vascular progenitors reside within adult blood vessels, making the small embryonic stem cell hypothesis less tenable.
Mesenchymal stem cells (MSCs) derived from adult bone marrow have been widely studied for their regenerative potential due to their ability to differentiate into various mesodermal lineages, including osteogenic, chondrogenic, adipogenic and perivascular lineages. However, despite extensive research, there is no strong evidence to support the notion that adult bone marrow-derived MSCs can differentiate into fully functional endothelial cells capable of participating in vasculogenesis or angiogenesis in vivo [29, 30]. In contrast, MSCs with a more immature phenotype, such as those derived from perinatal tissues, seem to exhibit a greater degree of plasticity and appear to have a broader differentiation potential. Wharton’s jelly-derived MSCs, which are obtained from the umbilical cord, have been shown to express both mesenchymal and endothelial markers more readily compared to their adult bone marrow counterparts. These cells can be induced to differentiate into endothelial-like cells [31, 32]. Their perinatal origin suggests that they retain an epigenetic profile that is more permissive to endothelial differentiation compared to adult MSCs, which may have undergone further lineage restriction. Another subset of MSCs with endothelial differentiation potential includes those isolated from CD133 + progenitor populations [19]. Furthermore, MSC-like cells derived from vascular tumors such as infantile hemangiomas have demonstrated the ability to undergo endothelial differentiation [33, 34]. These findings suggest that MSCs originating from sources that retain a closer developmental proximity to vascular progenitors may be more amenable to endothelial differentiation compared to bone marrow-derived MSCs. Gil et al. [35] demonstrate that mesodermal cell from induced pluripotent stem cells (iPSCs) can differentiate into ECFC-like cells, restoring vascular function in diabetic mice. Moreover, Pal et al. [36] introduce an entirely different model, proposing that fibroblasts can transdifferentiate into endothelial-like cells following injury. Using single-cell RNA sequencing, they found that fibroblasts reprogram into endothelial-like cells after miR-200b inhibition. This vasculogenic fibroblast subset contributes to wound healing and neovascularization. This challenges the assumption that ECFCs must derive from endothelial precursors, as non-endothelial cells may contribute to endothelial lineage plasticity. Their findings indicate that ECFCs may emerge from non-traditional sources, further complicating the assumption that they originate solely from small embryonic-like stem cells.
ECFC characterization
The characterization of ECFCs requires precise criteria to ensure reproducibility and consistency as previously described [3, 4]. These colonies should present a well-defined structure with a clonogenic nature, displaying a cobblestone morphology and comprising more than fifty adherent cells. Their abundance is quantified as the number of colonies per ten million mononuclear cells seeded, providing a standardized metric for comparison. Isolation efficiency is a crucial parameter, with expert laboratories achieving a success rate of seventy to seventy-five% in deriving these cells from the peripheral blood of healthy individuals. However, variability in donor samples must be acknowledged, as certain cases result in the absence of ECFC colonies. This outcome, referred to as “zero colonies,” is an essential data point that must be documented. Beyond initial isolation, the expansion potential of these cells should be assessed through both passage number and population doubling time, offering deeper insights into their proliferative capacity. Additionally, studies should systematically report the duration of culture, starting from the day of the original mononuclear cell isolation, known as day zero. By adhering to these rigorous characterization standards, research on ECFCs can maintain a high level of reliability, paving the way for advancements in vascular biology and regenerative medicine.
ECFCs are characterized by the presence of specific endothelial markers, including CD31, CD34 [37, 38], intracellular CD133 [39], VE-Cadherin, vascular endothelial growth factor receptor 2 (KDR), von Willebrand factor (VWF) in Weibel Palade bodies and MCAM (CD146) [40]. They also express Aldehyde Dehydrogenase (ALDH) [41,42,43]. ALDH is a group of enzymes that plays a crucial role in detoxifying aldehydes thus inducing protection against oxidative stress. ALDH is also involved in retinoic acid biosynthesis from retinaldehyde, which is crucial for cellular differentiation, and immune function and has an important role in maintaining “stemness,” the stem cells ability to self-renew and differentiate [44]. As shown in Fig. 1, ALDH is expressed by CB-ECFCs, and its expression decreases along in vitro expansion. This novel observation of ALDH expression and its decline with in vitro expansion adds a new dimension to understanding the biology of ECFCs. It suggests a potential link between ALDH activity and the maintenance of stemness or functional capabilities in ECFCs. This finding could provide valuable insights into optimizing culture conditions for therapeutic applications, preserving the regenerative potential of ECFCs, and further exploring the role of ALDH as a marker for stemness and cellular functionality. Finally, an overexpression of neuregulin-1 (NRG1) in ECFCs compared to mature endothelial cells has also been described [45]. Unlike classic endothelial cells, ECFCs actively secrete bioactive NRG1, promoting cardiomyocyte survival, proliferation, and engraftment through the PI3K/Akt signaling pathway. This effect is absent in mature endothelial cells. In co-culture experiments and in vivo models, ECFCs enhanced cardiomyocyte resistance to apoptosis and improved cardiac cell retention, whereas mature endothelial cells failed to provide similar benefits. Silencing NRG1 in ECFCs abolished these protective effects, confirming its essential role. This study establishes ECFCs as distinct from mature endothelial cells due to their regenerative potential, making them promising candidates for cardiovascular therapy, particularly in cardiac repair and ischemic heart disease treatment.

Aldehyde dehydrogenase (ALDH) activity in CB-ECFCs during in vitro expansion. Quantification of ALDH activity. ALDH activity was measured using the Aldefluor reagent (StemCell Technologies) following the manufacturer’s instructions. A concentration of 0.625 µg/mL Aldefluor substrate was added to 1 million CB-ECFCs in Aldefluor assay buffer and incubated for 30 min at 37 °C, converting the substrate to a fluorescent product. To ensure specificity, a portion of the Aldefluor-stained cells was treated with 5 µL of 1.5 mM diethylaminobenzaldehyde (DEAB), a specific ALDH inhibitor, as a control (center panel). After Aldefluor treatment, the cells were stained with fluorochrome-conjugated antibodies and the fluorescence was quantified using an Accuri C6 PLUS (BD Biosciences) (right panel). B:Evolution of ALDH activity of CB-ECFCs. CB-ECFCs were cultured in endothelial growth medium 2 (EGM2, Lonza Paris, France) and the quantification of ALDH activity was performed from 20 days after starting of the culture for several weeks. A significant decrease of ALDH-positive cells proportion was observed along culture and passages in culture. The x-axis represents the number of days after MNC culture
ECFC properties
In addition to the difference between the number of colonies obtained [46], CB-ECFCs exhibit superior proliferative capacity compared to their adult blood-derived counterparts [5, 47, 48] and demonstrate a remarkable resistance to apoptosis and stemness marker expression, distinguishing them from HUVECs [12, 39, 49], probably linked to, at least in part, longer telomeres [50]. CB-ECFCs have been observed to promote vessel formation [51], even better than human Embryonic Stem Cell-Endothelial Cells (hESC-EC) [52].
One of their key mechanisms of action is their direct integration into host tissues, where they actively participate in vessel formation, a capability that sets them apart from hematopoietic EPCs [5] and bone marrow-derived mesenchymal stem cells [30]. Unlike these other cell types, ECFCs can engraft directly into tissues and contribute to the formation of human vasculature in murine models of hind limb ischemia, underscoring their therapeutic potential in vascular regeneration [5, 10, 40]. Beyond direct incorporation, another crucial mechanism underlying ECFC functionality is their paracrine influence. Initially, ECFCs were believed to lack significant secretory activity, particularly in comparison to hematopoietic EPCs, as they produced neither vascular endothelial growth factor-A nor substantial levels of interleukin-8 [10]. However, further research has revealed that under inflammatory, stress, or senescent conditions, ECFCs possess a dynamic secretion profile, releasing interleukin-8 and other chemokines [53, 54], such as CXCL and CCL molecules [55], to actively recruit immune cells. Additionally, they facilitate perivascular cell recruitment through the secretion of platelet-derived growth factor-BB [56], thereby enhancing cellular engraftment and tissue integration. Their secretory capacity extends to the release of microvesicles and exosomes, which are believed to contribute significantly to their regenerative effects [57,58,59]. Another pivotal mechanism involves the direct supportive role ECFCs play in mesenchymal stem cell function. These interactions, observed across various mesenchymal stem and progenitor cell origins, suggest that ECFCs promote the differentiation of mesenchymal precursors into bona fide perivascular cells through a Notch/Jagged-1 signaling pathway [60]. This crosstalk is crucial for optimizing vascular stabilization and regeneration in ischemic conditions. Lastly, ECFCs exhibit a unique adhesive capacity that facilitates perivascular cell attachment via an endoglin-dependent mechanism [61]. When mesenchymal stem cells are co-administered in models of hind limb ischemia, endoglin binding by ECFCs accelerates perivascular cell engraftment, leading to enhanced vascular repair and expedited recovery [61]. This adhesive interaction represents a critical feature of ECFC-mediated vascular regeneration, further solidifying their role as a cornerstone in vascular therapeutic strategies. Nonetheless, numerous preclinical studies on vasculogenesis using human ECFCs have shown that for successful engraftment, it is often necessary to co-transplant perivascular support cells, such as mesenchymal stromal cells (MSCs) [29, 61,62,63]. Given the challenges of using multiple cell types in clinical settings, such as the complexity and regulatory hurdles associated with ECFCs and MSCs, alternative approaches are necessary. Lin and colleagues offer a promising solution by demonstrating the pivotal role of mitochondrial transfer from MSCs to endothelial cells (ECs) via tunneling nanotubes (TNTs). This mitochondrial transfer is crucial for the successful integration of ECFCs, suggesting that replacing MSCs with isolated mitochondria might simplify the clinical application while retaining the therapeutic benefits [64, 65].
Beyond vascular regeneration, the immunomodulatory properties of ECFCs
The use of induced Pluripotent Stem-derived endothelial cells (iPS-ECs) in regenerative medicine offers significant advantages, including the ability to engineer them to match the genetic profile of the patient and thus minimize the risk of immune rejection [2]. However, ECFCs, particularly those derived from CB exhibit a low immunogenicity level, in addition to their increased vasculogenic potential, making them highly valuable in this context. Indeed, the first description of EPC-derived ECs in 2010 demonstrated low levels of major histocompatibility complex (MHC) class I (MHC I) and no constitutive expression of MHC class II (MHC II) in contrast to mature ECs in rats [66]. MHC I and MHC II are responsible for distinct immune responses. MHC I expression on allogeneic tissue is responsible for the activation of cytotoxic CD8+ T cells [67] and complement-mediated lysis upon alloantibodies binding [68] while antigen recognition via MHC II is critical for the activation of allogeneic CD4+ T cell and the initiation of the direct pathway of allorecognition [69].
Interestingly, endothelial cells derived from endothelial progenitor cells (EPC-derived ECs) in rats exhibit low expression of MHC class I molecules. This low expression is linked to a reduced susceptibility to attack by pre-activated allospecific cytotoxic CD8 + T lymphocytes and complement-mediated lysis [66]. Moreover, upon stimulation with interferon-gamma (IFN-γ), EPC-derived ECs upregulated MHC I but marginally upregulated MHC II compared to mature ECs [66]. Therefore, ECFCs low expression of MHC molecules makes them less likely to be recognized and attacked by the immune system and limits their ability to activate the adaptive immune system through the antigen presentation pathway. Indeed, despite their allogeneic origin, when EPC-derived ECs were seeded into acellular aortic grafts and transplanted into rats, the grafts exhibited excellent endothelialization and induced mild inflammatory responses without signs of rejection [66]. This suggests that EPC-derived ECs can be used effectively in vascular reconstruction without eliciting a strong immune response.
The immune function of mature endothelial cells has been clearly established, notably their capacity to recognize and respond to stimuli such as proinflammatory cytokines IL-6, IL-1β and tumor necrosis factor alpha (TNF-α), but also pathogen-associated molecular patterns, through the expression of different Toll-like receptors (TLRs) [70]. ECFCs have been shown to inherently express the full spectrum of TLRs, ranging from TLR1 to TLR10, but data on their functional role in ECFC are still underexplored [71]. TLR3 recognizes double-stranded RNA (dsRNA), a molecule associated with viral infections and cellular stress, and its stimulation in CB-ECFCs induces the release of pro-inflammatory cytokines including TNF-α, IL-6, and IL-8, and the expression of ICAM-1 and VCAM-1 adhesion molecules [72]. Additionally, TLR3 stimulation in CB-ECFCs impairs their proliferation and angiogenic capacity, delaying ischemia recovery in a hind-limb ischemia mouse model [72]. This effect has been corroborated in an apolipoprotein E-deficient model of atherogenesis where TLR3 stimulation in cultured endothelial progenitor cells reduced their migration capacity while increasing the apoptosis rate and their production of reactive oxygen species. Importantly, the adoptive transfer of TLR3-stimulated ECFCs hampered reendothelialization in a model of artery injury, further confirming the role of TLR3 in endothelial dysfunction [73].
In contrast, stimulation with lipopolysaccharide (LPS) upregulated TLR2 mRNA expression and increased IL-6 production in isolated CB-ECFCs, but it did not impact their growth or differentiation. Interestingly, TLR2 has been implicated in mobilizing endothelial progenitors from the bone marrow into circulation in a murine model of periodontal infection [74]. Furthermore, TLR2 signaling appears to play a critical role in tissue regeneration by modulating angiogenesis and oxidative stress during inflammatory conditions [75]. These findings highlight the distinct roles of TLR3 and TLR2 in endothelial progenitor cell biology and vascular function. TLR3 exerts anti-angiogenic effects and may impede regenerative processes, whereas TLR2 appears to facilitate regenerative outcomes through the modulation of inflammation and angiogenesis. Collectively, these data indicate that TLRs, particularly TLRs 2, 3 and 4 are functionally active in ECFCs and can influence angiogenic processes and the response to inflammatory stimuli. Further studies are needed to decipher the specific roles of TLRs subsets in ECFCs, particularly how they modulate angiogenic function and respond to various microenvironmental factors, including inflammation, infections and allogeneic conditions. Cell surface markers, such as CD34, play a key role in cell recognition, immune regulation, and intercellular communication [76].
CD34, a well-known surface marker associated with hematopoietic and endothelial progenitor cells, has been increasingly recognized for its functional relevance in modulating endothelial and immune cell behavior [7]. A recent study published in 2023 investigated the role of CD34 expression on ECFCs and endothelial cells and its impact on immune cell responses [77]. Using transcriptomic and functional analysis, the authors showed that CD34− high ECFCs exhibited a lower proliferation rate, but higher secretion levels of IL-33 and Angiopoietin 2 compared to CD34low ECFCs. Angiopoietin is a proangiogenic cytokine that drives vascular remodeling through effects on Tie2 receptor signaling in endothelial cells [78]. Interestingly, it has been shown to promote regulatory T cells (Treg) expansion through the stimulation of TIE2 expressing monocytes [79]. The co-culture of CD34 high ECFCs with naïve peripheral blood mononuclear cells (PBMC) strongly induced the differentiation of Tregs through the IL-33/ST2 axis [77]. IL-33 is produced in response to tissue injury and inflammation [80] and has been recently shown to induce Tregs expansion that controls the inflammatory response without altering the adaptive immune cells’ function [81]. In conclusion, the ability of ECFCs CD34+ to produce IL-33 and Angiopoietin 2 highlights their potential role in promoting immune tolerance in response to tissue injury and inflammation.
The activation of the TNF-α/TNFR2 signaling pathway has also been recently described as an additional mechanism involved in the immunoregulatory function of ECFCs [82]. TNF receptor type II (TNFR2) is preferentially expressed by immunosuppressive cells, including Tregs, and myeloid-derived suppressor cells (MDSCs).Its activation plays an important role in attenuating pro-inflammatory responses and promoting tissue regeneration [83]. Importantly, priming ECFCs with an appropriate dose of TNF-α could effectively upregulate TNFR2 expression and their immunoregulatory function without an excessive increase of the pro-inflammatory TNFR1 subset, indicating that the ECFCs respond to the inflammatory milieu to subsequently control inflammation-related tissue injury [84]. Of note, the immunosuppressive effect of ECFCs is independent of Tregs, as they did not increase Foxp3 expression in conventional T cells [82]. This observation is aligned with the lack of IL-10 and TGFβ production by T cells following co-culture with either CB-ECFCs or AB-ECFCs [82].
In addition to promoting immunoregulatory pathways, further studies have shown that human CB-ECFCs significantly express less pro-inflammatory mediators, such as arachidonate 5-lipo-oxygenase (ALOX5), TNF-α, colony-stimulating factor-2 (CSF2) and IL1-β, than adults ECFCs [85]. Moreover, CB-ECFCs display a reduced capacity to activate allogeneic mononuclear cells as reflected by reduced mRNA expression of IL6, IL8, CSF2 and ICOSL compared to the one triggered by AB-ECFCs or mesenchymal cells. Surprisingly, CB-ECFCs exhibit a similar pattern of expression of HLA-ABC and HLA-DR antigens as mesenchymal stromal cells (MSCs), which are commonly transplanted irrespective of HLA-matching [85]. The concomitant expression of immunoregulatory molecules on CB-ECFCs such as HLA-G, a hallmark of cord blood-derived cells [86], may participate to their low immunogenicity. Indeed, this concept has been corroborated by the findings of Proust et al. who demonstrated that CB-ECFCs significantly inhibit PBMC allo-proliferation, unlike the HLA-II + lymphoblastoid cell line [87]. Mechanistically, CB-ECFCs highly expressed the immunosuppressive markers HLA-G, IL-10, and TGF-β1, allowing them to be tolerated and to promote a functional vascular network in immunocompetent mice. IL-10 has been shown to increase HLA-G mRNA levels in trophoblast cells [88] which promotes foeto-maternal tolerance by inhibiting maternal NK cells activation without impairing their antiviral immune function [89].
Altogether, these aforementioned immunomodulatory properties may explain the safety of ECFCs transplantation in several immunocompetent experimental animal models, without being rejected [90,91,92].
In addition to their immunoregulatory function, a recent study demonstrated that ECFCs exhibit adaptive-like immune features and can establish inflammatory memory upon initial exposure to the viral-like ligand Poly(I: C) that stimulates TLR3 pathway [93]. A secondary stimulation of pre-exposed Poly(I: C) ECFCs induced robust changes in DNA methylation patterns at promoter genes related to inflammatory and immunometabolism pathways, a reminiscent observation of the trained immunity phenomenon [94, 95]. Moreover, the ECFCs harbored a distinct enrichment of the E74-like factor 1 (ELF1), a transcription factor that regulates the antiviral immune response and is distinct from the type I interferon signaling pathway [96]. However, further investigations are required to show the functional translation of the remodeled transcriptome and epigenome following Poly(I: C) pre-exposure of the ECFCs. A crosstalk has been described between the epigenetic landscape and the differential expression of microRNA (miRNA) cellular levels through the regulation of DNA methylation modifications [97]. Although Grelier et al. observed upregulation of miR-29b, miR-146a, and miR-155 expression following Poly(I: C) stimulation of ECFCs, selectively inhibiting these microRNAs did not alter the cells’ phenotype or function [72]. This reflects the complexity of ECFC biology at the transcriptomic and functional levels and highlights the need for future studies to decipher the molecular network involved in their homeostasis. Collectively, these data highlight the immunomodulatory functions of ECFCs (Fig. 2) and suggest that in addition to their vasculogenic capacity, these cells can closely interact will the immune microenvironment to elicit a tolerogenic state, a feature that can be affected by inflammatory stimuli.

Immunomodulatory properties of ECFCs. ECFCs, especially those derived from cord blood, demonstrate low immunogenicity, characterized by reduced expression of MHC class I and II molecules, thereby minimizing immune recognition and rejection. This feature is enhanced by their capacity to express immunoregulatory molecules such as IL-10, IL-33, and HLA-G, contributing to their ability to promote immune tolerance and facilitate tissue repair. These cells express a variety of Toll-like receptors (TLRs), particularly TLR2, TLR3, and TLR4, which influence angiogenic processes according to the antigenic ligand present in the inflammatory microenvironment. Additionally, the expression of TNFR2 further supports their role in modulating immune response to inflammatory stimuli
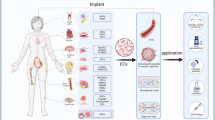
ECFCs: a promising approach for engineered organ revascularization (Fig. 3)

ECFCs are a promising approach for engineered organ revascularization. A Whole organ engineering consists of creating tridimensional acellular scaffolds for autologous or compatible cell seeing. The revascularization is the most critical step since it enables long-term survival of the seeded cells by providing blood flow through the whole scaffold. ECFCs hold promise for re-endothelialization of engineered organs through their multipotency, stemness and decreased immunogenicity. B Skin substitute prevascularization enhances integration within the host tissues. These engineered alternatives to skin grafts allow treating extended skin defects with low morbidity. Prevascularization demonstrated better integration within the recipient site. Tridimensional vascular networks could be obtained in vitro through ECFCs seeding
Tissue and organ engineering emerged in the 80’s as a promising alternative to allotransplantation. It aims to restore a function by building anatomical structures from acellular matrices and autologous or compatible cells. Several techniques exist to create tridimensional matrices (such as decellularization and 3D bioprinting) with encouraging results demonstrating the biocompatibility and absence of immunogenicity [98]. Growth factors such as Vascular Endothelial Growth Factor A (VEGF-A) and Epidermal Growth Factor (EGF) have been used to increase in vitro cell survival and multiplication within the scaffold, as well as mechanical stimulation such as shear forces and gravity. The progressive advances allowed great steps towards whole organ engineering, with promising results in liver, skin, lung and heart engineering. Moreover, the young field of Vascularized Composite Allotransplantation (VCA) has led to the development of reconstructive-aiming tissue engineering [99]. Building complex tissues would allow universal reconstructions in complex cases with no need for an immunosuppressive regimen, which is currently limiting VCA procedures. Multiple somatic cell seeding techniques have been described, leading to the successful colonization of tridimensional scaffolds [100]. However, to date, the main challenge for the prolonged success of these procedures is matrix revascularization. If simple contact and imbibition seem sufficient for thin acellular biostructures such as Acellular Dermal Matrices (ADMs), complex 3D scaffolds require parenchymal vascular tree restoration [101]. Similarly, novel engineered skin constructs are rapidly developing through multi-layer 3D bioprinting, increasing the need for adequate revascularization processes [102]. In decellularized scaffolds, authors have demonstrated the preservation of the vascular lumen, which could be used as an optimal bed for vessel regrowth. Still, no approach has demonstrated successful long-term results in sizeable scaffold re-endothelialization yet. Jank et al. demonstrated skin flap scaffold building with preservation of the vascular architecture, but pointed out the crucial importance of successful preparation of the vessels for re-endothelialization [103].
The evidence of using ECFCs for engineered construct revascularization remains poor to date. Cornerstone studies proved that ECFC seeding in collagen, hydrogels or matrigel implant [5, 29, 51, 62, 104, 105] were able to form functional blood vessels. Denecke et al. described in 2013 optimized ECFC culture on synthetic scaffold biomaterial, basing the culture media on human platelet lysate-enriched EGM-2 [106]. Their results showed minor endothelial tube formation and discrete adherence to the synthetic scaffold, but these encouraging results remain preliminary. It seems crucial to study evidence from alternative endothelial progenitors. Multiple attempts based on using Human Umbilical Vein Endothelial Cells (HUVECs) for acellular scaffold revascularization have been described. Hao et al. [107] experienced endothelial seeding using HUVECs in decellularized pancreatic scaffolds. While they showed a better potential for tube formation versus regular collagen scaffolds and demonstrated higher upregulation of pro-angiogenic factors such as MMP2, VEGF-A, and PAR-1 in endothelial cells cultured within decellularized scaffolds, they did not demonstrate 3D sprouting of vascular structures 24 h post-seeding. Watanabe et al. studied liver scaffold HUVEC-based revascularization and showed a partial coating of the portal vein sinusoids [108]. However, the endothelial cell phenotype was not studied despite being critical due to the sinusoid type found in the liver. Interestingly, they proposed using a perfusion machine to stimulate vessel tubulation through physiological-like shear forces. Despite this principle being highly promising, the lack of integrity of the endothelium lining led to major leakage that could affect the scaffold architecture [108]. Shaheen et al. produced pioneering results with successful perfusion-revascularization of full-sized liver scaffolds using HUVECs, with sinusoid-like characteristics [109]. Their technology was tested in vivo in an auxiliary allotransplantation model, showing endothelial immunogenicity. However, the perfusion and histological analysis of the reimplanted bioengineered livers showed a decreasing perfused and total graft volume, probably due to delayed thrombosis within the re-endothelialized vessels. The graft failure was significantly delayed by immunosuppressive drugs, emphasizing the importance of the immune reaction on the endothelial function. These results suggest the potential of HUVECs as a source for whole organ scaffold revascularization but highlight several limitations. Similarly, Yuan et al. used cultured Pulmonary Microvascular Endothelial Cells (PMECs) to achieve lung scaffold re-endothelialization [110]. They promoted cell migration and retention using a ROCK inhibitor to reduce cell size and facilitate intracapillary migration and used a machine-perfusion powered bioreactor to achieve intraluminal culture. Advanced TEM demonstrated lumen patency and endothelial coating, but the absence of type 1 alveolar epithelial cells and supporting vascular cells, such as pericytes and smooth muscle cells, prevented the full reconstruction of lung barrier functions. Although endothelial cells partially regained native phenotypes, their profiles remained incomplete, particularly for capillary subtypes. Their findings also missed key factors such as cyclic mechanical stretch, oxygen gradients, and paracrine signals, limiting biomimicry. Luo et al. achieved vascular construct re-endothelialization using a similar shear-stress enhanced bioreactor-based culture to enhance hiPSC-ECs alignment and adhesion [111]. Their engineered constructs demonstrated in vivo persistent patency and thrombosis prevention compared to non-endothelialized controls. These results were confirmed by Park et al. in a 2025 study using hiPSC-ECs and similar seeding techniques, with early evidence of smooth muscle recellularization and maturation [112].
The alternative of using stem/progenitor cells as a source of engineered organ revascularization potentially presents multiple advantages. Besides a higher potential of proliferation, which appears necessary due to the complex structure of a whole organ vascular tree, stem/progenitor cells can also lead to recreating the vascular microenvironment with supporting cell differentiation [113, 114]. Zhang et al. used a combination of human Adipose Stromal Cells (hASCs) and HUVECs for flap scaffold revascularization, which allowed successful microvascular implantation in nude rats [115]. They suggest the potential of hASCs towards an M2 macrophage phenotype, promoting an anti-inflammatory and immunotolerant environment. A recent study by Jiang et al. demonstrated successful 3D-printed scaffold revascularization using ESCs and ESC-derived vascular progenitor cells, with optimal integration to the recipient’s vasculature [116]. However, both their studies used nude rodent models and small constructs, limiting the interpretation to whole organ engineering with potential immunogenicity of ASC and ESC-derived vascular structures.
Using ECFCs in similar applications could significantly enhance vascular network formation within complex three-dimensional organ scaffolds suitable for clinical implantation. Indeed, their superior angiogenic potential, stable phenotype, and reduced immunogenicity would represent an asset. Specifically, CB-ECFCs exhibit greater angiogenic properties compared to HUVECs, endothelial cells derived from human embryonic stem cells or AB-ECFCs [5, 47,48,49, 52]. In artificial skin development, two main strategies prevail: the first involves artificially recreating skin using 2D/3D cell cultures, with or without 3D printing, and then implanting the bioengineered skin into the body to integrate with host tissue and promote vascularization. To attend to this objective, on the basis of previously skin organoids [117, 118], we developed a new biomaterial comprising a porous resorbable matrix [119] able to receive several cell subtypes. The porosity allows the material to have a structure similar to natural tissues, promoting cellular growth and integration when implanted into the body. This material will gradually dissolve or be absorbed by the surrounding skin over time, eliminating the need for a second surgery to remove it. The second strategy is decellularizing fresh skin for in vivo recellularization. In both approaches, enhancing the vascularization is crucial to ensure optimal engraftment. The use of ECFCs for simpler constructs, such as skin substitute pre-vascularization, has revealed successful blood vessel integration with enhanced healing [120]. If the challenge of skin substitute revascularization seems more affordable due to its decreased thickness and layered structure, this growing field is of crucial importance for addressing extended wounds as found in severe burns. ECFCs present a robust alternative for achieving enhanced outcomes through successful vascular tube formation and improved integration during the process of vascular self-sufficiency [121]. They demonstrate higher survival and proliferation potential [12, 47,48,49] and vasculogenic capability in vivo [51], along with immune privileges described here and stemness properties [39, 122, 123], making them an ideal cell type for pre vascularized organs or scaffolds.
Conclusion
All in all, this review highlights the unique vasculogenic properties and therapeutic potential of CB-ECFCs in regenerative medicine. ECFCs from cord blood exhibit significant advantages over other sources, including higher proliferative capacity, robust vasculogenic potential, and fewer ethical concerns compared to embryonic stem cells. Additionally, these cells demonstrate immune-privileged characteristics, reducing the likelihood of immune rejection and improving the success rates of allogenic cell therapies. These properties result from a dampened proinflammatory profile, higher immunoregulatory capabilities, and an enhanced capacity to retain memory from previous antigenic exposures compared to mature or adult endothelial cells. Thus CB-ECFCs represent a promising avenue for cell-based therapies, offering a combination of vasculogenic potential, immune privilege, and therapeutic versatility. Transcriptomic, phenotypic and functional studies are warranted to provide a comprehensive analysis of the immunological features of CB-ECFCs. Moreover, future research and clinical applications should focus on harnessing these vasculogenic and immunomodulatory properties to develop effective treatments for cardiovascular conditions, but also on developing pre-vascularized organs.
Data availability
No datasets were generated or analysed during the current study.
References
Wang J, Metheny L (2023) Umbilical cord blood derived cellular therapy: advances in clinical development. Front Oncol 13:1167266. https://doi.org/10.3389/fonc.2023.1167266
Smadja DM (2019) Vasculogenic stem and progenitor cells in human: future cell therapy product or liquid biopsy for vascular disease. Adv Exp Med Biol 1201:215–237. https://doi.org/10.1007/978-3-030-31206-0_11
Smadja DM, Melero-Martin JM, Eikenboom J et al (2019) Standardization of methods to quantify and culture endothelial colony-forming cells derived from peripheral blood: position paper from the international society on thrombosis and haemostasis SSC. J Thromb Haemost 17:1190–1194. https://doi.org/10.1111/jth.14462
Blandinieres A, Randi AM, Paschalaki KE et al (2023) Results of an international survey about methods used to isolate human endothelial colony-forming cells (ECFCs): guidance from the scientific and standardization committee on vascular biology of the international society of thrombosis and hemostasis. J Thromb Haemost S 1538–7836(23):00498–00491. https://doi.org/10.1016/j.jtha.2023.06.014
Yoder MC, Mead LE, Prater D et al (2007) Redefining endothelial progenitor cells via clonal analysis and hematopoietic stem/progenitor cell principals. Blood 109:1801–1809. https://doi.org/10.1182/blood-2006-08-043471
Asahara T, Murohara T, Sullivan A et al (1997) Isolation of putative progenitor endothelial cells for angiogenesis. Science 275:964–967. https://doi.org/10.1126/science.275.5302.964
Hassanpour M, Salybekov AA, Kobayashi S, Asahara T (2023) CD34 positive cells as endothelial progenitor cells in biology and medicine. Front Cell Dev Biol 11:1128134. https://doi.org/10.3389/fcell.2023.1128134
Lin Y, Weisdorf DJ, Solovey A, Hebbel RP (2000) Origins of Circulating endothelial cells and endothelial outgrowth from blood. J Clin Invest 105:71–77. https://doi.org/10.1172/JCI8071
Fujisawa T, Tura-Ceide O, Hunter A et al (2019) Endothelial progenitor cells do not originate from the bone marrow. Circulation 140:1524–1526. https://doi.org/10.1161/CIRCULATIONAHA.119.042351
Hur J, Yoon C-H, Kim H-S et al (2004) Characterization of two types of endothelial progenitor cells and their different contributions to neovasculogenesis. Arterioscler Thromb Vasc Biol 24:288–293. https://doi.org/10.1161/01.ATV.0000114236.77009.06
Delorme B, Basire A, Gentile C et al (2005) Presence of endothelial progenitor cells, distinct from mature endothelial cells, within human CD146 + blood cells. Thromb Haemost 94:1270–1279. https://doi.org/10.1160/TH05-07-0499
Smadja DM, Bièche I, Uzan G et al (2005) PAR-1 activation on human late endothelial progenitor cells enhances angiogenesis in vitro with upregulation of the SDF-1/CXCR4 system. Arterioscler Thromb Vasc Biol 25:2321–2327. https://doi.org/10.1161/01.ATV.0000184762.63888.bd
Lee JH, Lee SH, Yoo SY et al (2013) CD34 hybrid cells promote endothelial Colony-Forming cell bioactivity and therapeutic potential for ischemic diseases. ATVB 33:1622–1634. https://doi.org/10.1161/ATVBAHA.112.301052
Case J, Mead LE, Bessler WK et al (2007) Human CD34 + AC133 + VEGFR-2 + cells are not endothelial progenitor cells but distinct, primitive hematopoietic progenitors. Exp Hematol 35:1109–1118. https://doi.org/10.1016/j.exphem.2007.04.002
Timmermans F, Van Hauwermeiren F, De Smedt M et al (2007) Endothelial outgrowth cells are not derived from CD133 + cells or CD45 + hematopoietic precursors. Arterioscler Thromb Vasc Biol 27:1572–1579. https://doi.org/10.1161/ATVBAHA.107.144972
Tura O, Skinner EM, Barclay GR et al (2013) Late outgrowth endothelial cells resemble mature endothelial cells and are not derived from bone marrow. Stem Cells 31:338–348. https://doi.org/10.1002/stem.1280
Lin Y, Banno K, Gil C-H et al (2023) Origin, prospective identification, and function of Circulating endothelial colony-forming cells in mice and humans. JCI Insight 8:e164781. https://doi.org/10.1172/jci.insight.164781
Ratajczak MZ, Ratajczak J, Kucia M (2019) Very small Embryonic-Like stem cells (VSELs). Circ Res 124:208–210. https://doi.org/10.1161/CIRCRESAHA.118.314287
Guerin CL, Loyer X, Vilar J et al (2015) Bone-marrow-derived very small embryonic-like stem cells in patients with critical leg ischaemia: evidence of vasculogenic potential. Thromb Haemost 113:1084–1094. https://doi.org/10.1160/TH14-09-0748
Lahlil R, Scrofani M, Barbet R et al (2018) VSELs maintain their pluripotency and competence to differentiate after enhanced ex vivo expansion. Stem Cell Rev Rep 14:510–524. https://doi.org/10.1007/s12015-018-9821-1
Domingues A, Rossi E, Bujko K et al (2022) Human CD34 + very small embryonic-like stem cells can give rise to endothelial colony-forming cells with a multistep differentiation strategy using UM171 and nicotinamide acid. https://doi.org/10.1038/s41375-022-01517-0. Leukemia
Saucourt C, Vogt S, Merlin A et al (2019) Design and validation of an automated process for the expansion of peripheral Blood-Derived CD34 + Cells for clinical use after myocardial infarction. Stem Cells Transl Med 8:822–832. https://doi.org/10.1002/sctm.17-0277
Hénon P, Kowalczyk M, Aries A et al (2022) Industrialized GMP production of CD34 + Cells (ProtheraCytes®) at clinical scale for treatment of ischemic cardiac diseases is feasible and safe. Stem Cell Rev Rep 18:1614–1626. https://doi.org/10.1007/s12015-022-10373-5
Hénon P (2020) Key success factors for regenerative medicine in acquired heart diseases. Stem Cell Rev Rep 16:441–458. https://doi.org/10.1007/s12015-020-09961-0
Wakabayashi T, Naito H, Suehiro J-I et al (2018) CD157 marks Tissue-Resident endothelial stem cells with homeostatic and regenerative properties. Cell Stem Cell 22:384–397e6. https://doi.org/10.1016/j.stem.2018.01.010
Naito H, Wakabayashi T, Ishida M et al (2020) Isolation of tissue-resident vascular endothelial stem cells from mouse liver. Nat Protoc 15:1066–1081. https://doi.org/10.1038/s41596-019-0276-x
Bhagwani AR, Farkas D, Harmon B et al (2020) Clonally selected primitive endothelial cells promote occlusive pulmonary arteriopathy and severe pulmonary hypertension in rats exposed to chronic hypoxia. Sci Rep 10:1136. https://doi.org/10.1038/s41598-020-58083-7
Lin Y, Gil C-H, Banno K et al (2024) ABCG2-Expressing clonal repopulating endothelial cells serve to form and maintain blood vessels. Circulation 150:451–465. https://doi.org/10.1161/CIRCULATIONAHA.122.061833
Melero-Martin JM, De Obaldia ME, Kang S-Y et al (2008) Engineering robust and functional vascular networks in vivo with human adult and cord blood-derived progenitor cells. Circ Res 103:194–202. https://doi.org/10.1161/CIRCRESAHA.108.178590
Smadja DM, d’Audigier C, Guerin CL et al (2012) Angiogenic potential of BM MSCs derived from patients with critical leg ischemia. Bone Marrow Transpl 47:997–1000. https://doi.org/10.1038/bmt.2011.196
Wang S-H, Lin S-J, Chen Y-H et al (2009) Late outgrowth endothelial cells derived from wharton jelly in human umbilical cord reduce neointimal formation after vascular injury: involvement of pigment epithelium-derived factor. Arterioscler Thromb Vasc Biol 29:816–822. https://doi.org/10.1161/ATVBAHA.109.184739
Aguilera V, Briceño L, Contreras H et al (2014) Endothelium trans differentiated from Wharton’s jelly mesenchymal cells promote tissue regeneration: potential role of soluble pro-angiogenic factors. PLoS ONE 9:e111025. https://doi.org/10.1371/journal.pone.0111025
Khan ZA, Boscolo E, Picard A et al (2008) Multipotential stem cells recapitulate human infantile hemangioma in immunodeficient mice. J Clin Invest 118:2592–2599. https://doi.org/10.1172/JCI33493
Smadja DM, Guerin CL, Boscolo E et al (2014) α6-Integrin is required for the adhesion and vasculogenic potential of hemangioma stem cells. Stem Cells 32:684–693. https://doi.org/10.1002/stem.1539
Gil C-H, Chakraborty D, Vieira CP et al (2022) Specific mesoderm subset derived from human pluripotent stem cells ameliorates microvascular pathology in type 2 diabetic mice. Sci Adv 8:eabm5559. https://doi.org/10.1126/sciadv.abm5559
Pal D, Ghatak S, Singh K et al (2023) Identification of a physiologic vasculogenic fibroblast state to achieve tissue repair. Nat Commun 14:1129. https://doi.org/10.1038/s41467-023-36665-z
Tasev D, Konijnenberg LSF, Amado-Azevedo J et al (2016) CD34 expression modulates tube-forming capacity and barrier properties of peripheral blood-derived endothelial colony-forming cells (ECFCs). Angiogenesis 19:325–338. https://doi.org/10.1007/s10456-016-9506-9
Sherman SE, Kuljanin M, Cooper TT et al (2020) Purification and functional characterization of CD34-Expressing cell subsets following ex vivo expansion of umbilical cord Blood-Derived endothelial Colony-Forming cells. Stem Cells Dev 29:895–910. https://doi.org/10.1089/scd.2020.0008
Rossi E, Poirault-Chassac S, Bieche I et al (2019) Human endothelial colony forming cells express intracellular CD133 that modulates their vasculogenic properties. Stem Cell Rev Rep 15:590–600. https://doi.org/10.1007/s12015-019-09881-8
Silvestre J-S, Smadja DM, Lévy BI (2013) Postischemic revascularization: from cellular and molecular mechanisms to clinical applications. Physiol Rev 93:1743–1802. https://doi.org/10.1152/physrev.00006.2013
Nagano M, Yamashita T, Hamada H et al (2007) Identification of functional endothelial progenitor cells suitable for the treatment of ischemic tissue using human umbilical cord blood. Blood 110:151–160. https://doi.org/10.1182/blood-2006-10-047092
Putman DM, Liu KY, Broughton HC et al (2012) Umbilical cord Blood-Derived aldehyde Dehydrogenase‐Expressing progenitor cells promote recovery from acute ischemic injury. Stem Cells 30:2248–2260. https://doi.org/10.1002/stem.1206
Povsic TJ, Zavodni KL, Kelly FL et al (2007) Circulating progenitor cells can be reliably identified on the basis of aldehyde dehydrogenase activity. J Am Coll Cardiol 50:2243–2248. https://doi.org/10.1016/j.jacc.2007.08.033
Mezquita B, Mezquita C (2019) Two opposing faces of retinoic acid: induction of stemness or induction of differentiation depending on Cell-Type. Biomolecules 9:567. https://doi.org/10.3390/biom9100567
Hong X, Oh N, Wang K et al (2021) Human endothelial colony-forming cells provide trophic support for pluripotent stem cell-derived cardiomyocytes via distinctively high expression of neuregulin-1. Angiogenesis 24:327–344. https://doi.org/10.1007/s10456-020-09765-3
Ingram DA, Mead LE, Tanaka H et al (2004) Identification of a novel hierarchy of endothelial progenitor cells using human peripheral and umbilical cord blood. Blood 104:2752–2760. https://doi.org/10.1182/blood-2004-04-1396
Smadja DM, Bièche I, Emmerich J et al (2006) PAR-1 activation has different effects on the angiogenic activity of endothelial progenitor cells derived from human adult and cord blood. J Thromb Haemost 4:2729–2731. https://doi.org/10.1111/j.1538-7836.2006.02208.x
Smadja DM, Mauge L, Susen S et al (2009) Blood outgrowth endothelial cells from cord blood and peripheral blood: angiogenesis-related characteristics in vitro: a rebuttal. J Thromb Haemost 7:504–506 author reply 506–508. https://doi.org/10.1111/j.1538-7836.2008.03247.x
He T, Peterson TE, Holmuhamedov EL et al (2004) Human endothelial progenitor cells tolerate oxidative stress due to intrinsically high expression of manganese superoxide dismutase. Arterioscler Thromb Vasc Biol 24:2021–2027. https://doi.org/10.1161/01.ATV.0000142810.27849.8f
Toupance S, Simonici S, Labat C et al (2021) Number and replating capacity of endothelial Colony-Forming cells are telomere length dependent: implication for human atherogenesis. J Am Heart Assoc 10:e020606. https://doi.org/10.1161/JAHA.120.020606
Au P, Duda DG, Cohen KS et al (2008) Differential in vivo potential of endothelial progenitor cells from human umbilical cord blood and adult peripheral blood to form functional long-lasting vessels. Blood 111:1302–1305. https://doi.org/10.1182/blood-2007-06-094318
Smadja DM, Mauge L, Rancic J et al (2024) Comparative evaluation of endothelial Colony-Forming cells from cord and adult blood vs. Human embryonic stem Cell-Derived endothelial cells: insights into therapeutic angiogenesis potential. Stem Cell Rev Rep. https://doi.org/10.1007/s12015-024-10830-3
Smadja DM, Bièche I, Susen S et al (2009) Interleukin 8 is differently expressed and modulated by PAR-1 activation in early and late endothelial progenitor cells. J Cell Mol Med 13:2534–2546. https://doi.org/10.1111/j.1582-4934.2008.00429.x
Blandinières A, Gendron N, Bacha N et al (2019) Interleukin-8 release by endothelial colony-forming cells isolated from idiopathic pulmonary fibrosis patients might contribute to their pathogenicity. Angiogenesis 22:325–339. https://doi.org/10.1007/s10456-018-09659-5
d’Audigier C, Cochain C, Rossi E et al (2015) Thrombin receptor PAR-1 activation on endothelial progenitor cells enhances chemotaxis-associated genes expression and leukocyte recruitment by a COX-2-dependent mechanism. Angiogenesis 18:347–359. https://doi.org/10.1007/s10456-015-9471-8
Lin R-Z, Moreno-Luna R, Li D et al (2014) Human endothelial colony-forming cells serve as trophic mediators for mesenchymal stem cell engraftment via paracrine signaling. Proc Natl Acad Sci U S A 111:10137–10142. https://doi.org/10.1073/pnas.1405388111
Simoncini S, Chateau A-L, Robert S et al (2017) Biogenesis of Pro-senescent microparticles by endothelial colony forming cells from premature neonates is driven by SIRT1-Dependent epigenetic regulation of MKK6. Sci Rep 7:8277. https://doi.org/10.1038/s41598-017-08883-1
Bacha NC, Blandinieres A, Rossi E et al (2018) Endothelial microparticles are associated to pathogenesis of idiopathic pulmonary fibrosis. Stem Cell Rev Rep 14:223–235. https://doi.org/10.1007/s12015-017-9778-5
Dellett M, Brown ED, Guduric-Fuchs J et al (2017) MicroRNA-containing extracellular vesicles released from endothelial colony-forming cells modulate angiogenesis during ischaemic retinopathy. J Cell Mol Med 21:3405–3419. https://doi.org/10.1111/jcmm.13251
Boscolo E, Mulliken JB, Bischoff J (2011) VEGFR-1 mediates endothelial differentiation and formation of blood vessels in a murine model of infantile hemangioma. Am J Pathol 179:2266–2277. https://doi.org/10.1016/j.ajpath.2011.07.040
Rossi E, Smadja D, Goyard C et al (2017) Co-injection of mesenchymal stem cells with endothelial progenitor cells accelerates muscle recovery in Hind limb ischemia through an endoglin-dependent mechanism. Thromb Haemost 117:1908–1918. https://doi.org/10.1160/TH17-01-0007
Smadja DM, Levy M, Huang L et al (2015) Treprostinil indirectly regulates endothelial colony forming cell angiogenic properties by increasing VEGF-A produced by mesenchymal stem cells. Thromb Haemost 114:735–747. https://doi.org/10.1160/TH14-11-0907
Tripathi H, Domingues A, Donahue R et al (2023) Combined transplantation of human MSCs and ECFCs improves cardiac function and decrease cardiomyocyte apoptosis after acute myocardial infarction. Stem Cell Rev Rep 19:573–577. https://doi.org/10.1007/s12015-022-10468-z
Lin R-Z, Im G-B, Luo AC et al (2024) Mitochondrial transfer mediates endothelial cell engraftment through mitophagy. Nature 629:660–668. https://doi.org/10.1038/s41586-024-07340-0
Smadja DM (2024) Extracellular microvesicles vs. Mitochondria: competing for the top spot in cardiovascular regenerative medicine. Stem Cell Rev Rep. https://doi.org/10.1007/s12015-024-10758-8
Ladhoff J, Fleischer B, Hara Y et al (2010) Immune privilege of endothelial cells differentiated from endothelial progenitor cells. Cardiovascular Res 88:121–129. https://doi.org/10.1093/cvr/cvq109
Non-hematopoietic allograft cells directly activate CD8 + T cells and trigger acute rejection: An alternative mechanism of allorecognition| Nature Medicine. https://www.nature.com/articles/nm0302-233. Accessed 28 Aug 2024
Bian H, Reed EF (1999) Alloantibody-mediated class I signal transduction in endothelial cells and smooth muscle cells: enhancement by IFN-gamma and TNF-alpha. J Immunol 163:1010–1018
Leddon SA, Sant AJ (2010) Generation of MHC class II:peptide ligands for CD4 T cell allorecognition of MHC class II molecules. Curr Opin Organ Transpl 15:505–511. https://doi.org/10.1097/MOT.0b013e32833bfc5c
Immunomodulation by endothelial cells — partnering up with the immune system?| Nature Reviews Immunology. https://www.nature.com/articles/s41577-022-00694-4. Accessed 12 Aug 2024
Mazzucchelli I, Lisini D, Garofoli F et al (2015) Expression and function of toll-like receptors in human Circulating endothelial colony forming cells. Immunol Lett 168:98–104. https://doi.org/10.1016/j.imlet.2015.09.014
Grelier A, Cras A, Balitrand N et al (2013) Toll-like receptor 3 regulates cord blood-derived endothelial cell function in vitro and in vivo. Angiogenesis 16:821–836. https://doi.org/10.1007/s10456-013-9358-5
Zimmer S, Steinmetz M, Asdonk T et al (2011) Activation of endothelial Toll-Like receptor 3 impairs endothelial function. Circul Res 108:1358–1366. https://doi.org/10.1161/CIRCRESAHA.111.243246
Kebschull M, Haupt M, Jepsen S et al (2013) Mobilization of endothelial progenitors by recurrent bacteremias with a periodontal pathogen. PLoS ONE 8:e54860. https://doi.org/10.1371/journal.pone.0054860
West XZ, Malinin NL, Merkulova AA et al (2010) Oxidative stress induces angiogenesis by activating TLR2 with novel endogenous ligands. Nature 467:972–976. https://doi.org/10.1038/nature09421
Armingol E, Officer A, Harismendy O, Lewis NE (2021) Deciphering cell–cell interactions and communication from gene expression. Nat Rev Genet 22:71–88. https://doi.org/10.1038/s41576-020-00292-x
Arakelian L, Lion J, Churlaud G et al (2023) Endothelial CD34 expression and regulation of immune cell response in-vitro. Sci Rep 13:13512. https://doi.org/10.1038/s41598-023-40622-7
Eklund L, Olsen BR (2006) Tie receptors and their angiopoietin ligands are context-dependent regulators of vascular remodeling. Exp Cell Res 312:630–641. https://doi.org/10.1016/j.yexcr.2005.09.002
Coffelt SB, Chen Y-Y, Muthana M et al (2011) Angiopoietin 2 stimulates TIE2-Expressing monocytes to suppress T cell activation and to promote regulatory T cell expansion. J Immunol 186:4183–4190. https://doi.org/10.4049/jimmunol.1002802
Liew FY, Girard J-P, Turnquist HR (2016) Interleukin-33 in health and disease. Nat Rev Immunol 16:676–689. https://doi.org/10.1038/nri.2016.95
Faustino LD, Griffith JW, Rahimi RA et al (2020) Interleukin-33 activates regulatory T cells to suppress innate Γδ T cell responses in the lung. Nat Immunol 21:1371–1383. https://doi.org/10.1038/s41590-020-0785-3
Naserian S, Abdelgawad ME, Afshar Bakshloo M et al (2020) The TNF/TNFR2 signaling pathway is a key regulatory factor in endothelial progenitor cell immunosuppressive effect. Cell Commun Signal 18:94. https://doi.org/10.1186/s12964-020-00564-3
Chen Y, Jiang M, Chen X (2023) Therapeutic potential of TNFR2 agonists: a mechanistic perspective. Front Immunol 14:1209188. https://doi.org/10.3389/fimmu.2023.1209188
Nouri Barkestani M, Shamdani S, Afshar Bakshloo M et al (2021) TNFα priming through its interaction with TNFR2 enhances endothelial progenitor cell immunosuppressive effect: new hope for their widespread clinical application. Cell Commun Signal 19:1. https://doi.org/10.1186/s12964-020-00683-x
Nuzzolo ER, Capodimonti S, Martini M et al (2014) Adult and cord blood endothelial progenitor cells have different gene expression profiles and Immunogenic potential. Blood Transfus 12 Suppl 1s367–374. https://doi.org/10.2450/2013.0042-13
Buzzi M, Alviano F, Campioni D et al (2012) Umbilical cord blood CD34(+)cell-derived progeny produces human leukocyte antigen-G molecules with immuno-modulatory functions. Hum Immunol 73:150–155. https://doi.org/10.1016/j.humimm.2011.12.003
Proust R, Ponsen A-C, Rouffiac V et al (2020) Cord blood-endothelial colony forming cells are immunotolerated and participate at post-ischemic angiogenesis in an original dorsal chamber immunocompetent mouse model. Stem Cell Res Ther 11:172. https://doi.org/10.1186/s13287-020-01687-7
Moreau P, Adrian-Cabestre F, Menier C et al (1999) IL-10 selectively induces HLA-G expression in human trophoblasts and monocytes. Int Immunol 11:803–811. https://doi.org/10.1093/intimm/11.5.803
Tilburgs T, Evans JH, Crespo ÂC, Strominger JL (2015) The HLA-G cycle provides for both NK tolerance and immunity at the maternal–fetal interface. Proceedings of the National Academy of Sciences 112:13312–13317. https://doi.org/10.1073/pnas.1517724112
Bell A, Watt AP, Dudink I et al (2023) Endothelial colony forming cell administration promotes neurovascular unit development in growth restricted and appropriately grown fetal lambs. Stem Cell Res Ther 14:29. https://doi.org/10.1186/s13287-023-03249-z
Hubert L, Darbousset R, Panicot-Dubois L et al (2014) Neutrophils recruit and activate human endothelial colony-forming cells at the site of vessel injury via P-selectin glycoprotein ligand-1 and L-selectin. J Thromb Haemost 12:1170–1181. https://doi.org/10.1111/jth.12551
Flex A, Biscetti F, Iachininoto MG et al (2016) Human cord blood endothelial progenitors promote post-ischemic angiogenesis in immunocompetent mouse model. Thromb Res 141:106–111. https://doi.org/10.1016/j.thromres.2016.03.012
Weiss E, Vlahos A, Kim B et al (2021) Transcriptomic remodelling of fetal endothelial cells during establishment of inflammatory memory. Front Immunol 12:757393. https://doi.org/10.3389/fimmu.2021.757393
Jeljeli M, Riccio LGC, Doridot L et al (2019) Trained immunity modulates inflammation-induced fibrosis. Nat Commun 10:5670. https://doi.org/10.1038/s41467-019-13636-x
Jeljeli M, Riccio LGC, Chouzenoux S et al (2020) Macrophage immune memory controls endometriosis in mice and humans. Cell Rep 33:108325. https://doi.org/10.1016/j.celrep.2020.108325
Seifert LL, Si C, Saha D et al (2019) The ETS transcription factor ELF1 regulates a broadly antiviral program distinct from the type I interferon response. PLoS Pathog 15:e1007634. https://doi.org/10.1371/journal.ppat.1007634
Wang S, Wu W, Claret FX (2017) Mutual regulation of MicroRNAs and DNA methylation in human cancers. Epigenetics 12:187–197. https://doi.org/10.1080/15592294.2016.1273308
Berkane Y, Oubari H, van Dieren L et al (2024) Tissue engineering strategies for breast reconstruction: a literature review of current advances and future directions. Ann Transl Med 12:15. https://doi.org/10.21037/atm-23-1724
Lupon E, Lellouch AG, Acun A et al (2022) Engineering vascularized composite allografts using natural scaffolds: A systematic review. Tissue Eng Part B Rev 28:677–693. https://doi.org/10.1089/ten.TEB.2021.0102
Adil A, Xu M, Haykal S (2022) Recellularization of bioengineered scaffolds for vascular composite allotransplantation. Front Surg 9:843677. https://doi.org/10.3389/fsurg.2022.843677
Yu Y, Alkhawaji A, Ding Y, Mei J (2016) Decellularized scaffolds in regenerative medicine. Oncotarget 7:58671–58683. https://doi.org/10.18632/oncotarget.10945
Balavigneswaran CK, Selvaraj S, Vasudha TK et al (2023) Tissue engineered skin substitutes: A comprehensive review of basic design, fabrication using 3D printing, recent advances and challenges. Biomater Adv 153:213570. https://doi.org/10.1016/j.bioadv.2023.213570
Jank BJ, Goverman J, Guyette JP et al (2017) Creation of a bioengineered skin flap scaffold with a perfusable vascular pedicle. Tissue Eng Part A 23:696–707. https://doi.org/10.1089/ten.TEA.2016.0487
Allen P, Melero-Martin J, Bischoff J (2011) Type I collagen, fibrin and puramatrix matrices provide permissive environments for human endothelial and mesenchymal progenitor cells to form neovascular networks. J Tissue Eng Regen Med 5:e74–86. https://doi.org/10.1002/term.389
Allen P, Kang K-T, Bischoff J (2015) Rapid onset of perfused blood vessels after implantation of ECFCs and MPCs in collagen, puramatrix and fibrin provisional matrices. J Tissue Eng Regen Med 9:632–636. https://doi.org/10.1002/term.1803
Denecke B, Horsch LD, Radtke S et al (2015) Human endothelial colony-forming cells expanded with an improved protocol are a useful endothelial cell source for scaffold-based tissue engineering. J Tissue Eng Regen Med 9:E84–97. https://doi.org/10.1002/term.1673
Hao L, Khajouei F, Rodriguez J et al (2024) Unlocking the promise of decellularized pancreatic tissue: A novel approach to support angiogenesis in engineered tissue. Bioeng (Basel) 11:183. https://doi.org/10.3390/bioengineering11020183
Watanabe M, Yano K, Okawa K et al (2019) Construction of sinusoid-scale microvessels in perfusion culture of a decellularized liver. Acta Biomater 95:307–318. https://doi.org/10.1016/j.actbio.2018.12.042
Shaheen MF, Joo DJ, Ross JJ et al (2020) Sustained perfusion of revascularized bioengineered livers heterotopically transplanted into immunosuppressed pigs. Nat Biomed Eng 4:437–445. https://doi.org/10.1038/s41551-019-0460-x
Yuan Y, Leiby KL, Greaney AM et al (2021) A pulmonary vascular model from endothelialized whole organ scaffolds. Front Bioeng Biotechnol 9:760309. https://doi.org/10.3389/fbioe.2021.760309
Luo J, Qin L, Park J et al (2022) Readily available Tissue-Engineered vascular grafts derived from human induced pluripotent stem cells. Circ Res 130:925–927. https://doi.org/10.1161/CIRCRESAHA.121.320315
Park J, Riaz M, Qin L et al (2025) Fully biologic endothelialized-tissue-engineered vascular conduits provide antithrombotic function and graft patency. Cell Stem Cell 32:137–143e6. https://doi.org/10.1016/j.stem.2024.11.006
Gattazzo F, Urciuolo A, Bonaldo P (2014) Extracellular matrix: A dynamic microenvironment for stem cell niche. Biochimica et biophysica acta (BBA). - Gen Subj 1840:2506–2519. https://doi.org/10.1016/j.bbagen.2014.01.010
Burdick JA, Vunjak-Novakovic G (2009) Engineered microenvironments for controlled stem cell differentiation. Tissue Eng Part A 15:205–219. https://doi.org/10.1089/ten.tea.2008.0131
Zhang Q, Johnson JA, Dunne LW et al (2016) Decellularized skin/adipose tissue flap matrix for engineering vascularized composite soft tissue flaps. Acta Biomater 35:166–184. https://doi.org/10.1016/j.actbio.2016.02.017
Jiang W-C, Hsu W-Y, Ao-Ieong W-S et al (2021) A novel engineered vascular construct of stem cell-laden 3D-printed PGSA scaffold enhances tissue revascularization. Biofabrication 13. https://doi.org/10.1088/1758-5090/ac1259
Ramovs V, Janssen H, Fuentes I et al (2022) Characterization of the epidermal-dermal junction in hiPSC-derived skin organoids. Stem Cell Rep 17:1279–1288. https://doi.org/10.1016/j.stemcr.2022.04.008
Nasrallah A, Rezvani H-R, Kobaisi F et al (2024) Generation and characterization of CRISPR-Cas9-mediated XPC gene knockout in human skin cells. Sci Rep 14:30879. https://doi.org/10.1038/s41598-024-81675-6
Lellouch AG, Lantieri LA, Rachidi W, JOBEILI LM BIOMATERIAL COMPRISING A POROUS RESORBABLE MATRIX AND ASSOCIATED MANUFACTURING METHOD - WO 2021/198177 Al. Patent
Dai N-T, Huang W-S, Chang F-W et al (2018) Development of a novel Pre-Vascularized Three-Dimensional skin substitute using blood plasma gel. Cell Transpl 27:1535–1547. https://doi.org/10.1177/0963689718797570
Berkane Y, Kostyra DM, Chrelias T et al (2023) The autonomization principle in vascularized flaps: an alternative strategy for composite tissue scaffold in vivo revascularization. Bioeng (Basel) 10:1440. https://doi.org/10.3390/bioengineering10121440
Smadja DM, Bièche I, Silvestre J-S et al (2008) Bone morphogenetic proteins 2 and 4 are selectively expressed by late outgrowth endothelial progenitor cells and promote neoangiogenesis. Arterioscler Thromb Vasc Biol 28:2137–2143. https://doi.org/10.1161/ATVBAHA.108.168815
Smadja DM, Rossi E, Haviari S et al (2023) Thrombin receptor PAR1 Silencing in endothelial colony-forming cells modifies stemness and vasculogenic properties. J Thromb Haemost 21:3640–3648. https://doi.org/10.1016/j.jtha.2023.08.029
Funding
Open access funding provided by SCELC, Statewide California Electronic Library Consortium
This work was supported by the Priority Research Program and Equipment (PEPR) “Biotherapies and Bioproduction of Innovative Therapies” (BBTI), as a part of the government’s “France 2030” investment plan, through the program “Bioengineered Skin-France” (ANR-22-PEBI-0010). Cecile Pinault was supported by a grant of Domaine d’Intérêt Majeur (DIM) de la Région Île-de-France - BioConvergence pour la Santé (BIOCONVS).
Author information
Authors and Affiliations
Contributions
D. M. S. designed the present study, performed and/or analyzed the data, and wrote the manuscript. A. G. L. and M. J. also designed the present study and contributed to writing the manuscript. Y. B., N. K. B., J. R., A. C., C. P., M. V., E. P. and W. R. contributed to the manuscript by drafting specific sections and reviewing the text for intellectual content. All authors reviewed and approved the final manuscript.
Corresponding author
Ethics declarations
Ethical approval
Human umbilical cord were provided by Saint-Louis Hospital Biological Resources Center - Cord Blood Bank after information and consent of mothers, under French Health Ministry authorization (n°AC-2016-2759).
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Smadja, D.M., Berkane, Y., Bentounes, N.K. et al. Immune-privileged cord blood-derived endothelial colony-forming cells: advancing immunomodulation and vascular regeneration. Angiogenesis 28, 19 (2025). https://doi.org/10.1007/s10456-025-09973-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10456-025-09973-9