
Abstract
The Tonian Period (1000–720 Ma) bore witness to the transition from a prokaryote-dominated marine ecosystem to one characterized by the proliferation of eukaryotes. This fundamental shift has generally been attributed to evolving marine redox states. Here, we present sedimentological and geochemical analyses of the early Tonian Huainan, Feishui, and Huaibei groups in the Xuhuai basin of the North China craton. Multiple redox proxies show consistent, water depth-dependent variations across the Xuhuai basin. Excess barium contents and Ba/Al ratios further highlight spatial variations in primary productivity which ultimately regulate basinal redox structures. We propose that a shallow-water oxygen minimum zone sandwiched between the oxic/suboxic mid-depth and surface layer water masses occur in the oligotrophic Xuhuai basin, which is analogous to, but much shallower than modern oxygen minimum zones. Such marine redox architectures may benefit the maintenance of a bioavailable nitrate reservoir in the ocean, foreboding the subsequent expansion of eukaryotes.
Similar content being viewed by others

Introduction
The Tonian Period (1000–720 Ma) is a pivotal time for the evolution of eukaryotes. Increasing evidence indicates that the transition from prokaryote-dominated to eukaryote-dominated marine ecosystems did not happen until the Tonian Period1,2. Many revolutionary innovations of eukaryotes also occurred during this time, including the appearance of crown-group algae, biomineralization, and putative eukaryovory3. Broadly, eukaryotic diversification is associated with the ocean chemistry, particularly the concentrations of oxygen and bioavailable phosphorus and nitrate, which were expected to be deeply linked to the marine redox states4. However, the causal relationships between the eukaryotic evolutions and the ocean redox states remain elusive due to the disparate ocean redox conditions reconstructed using different redox proxies in various basins5,6,7,8,9,10.
The conventional model suggested that the ocean was stratified for most of the Proterozoic5. Beneath the oxygenated surface ocean, dynamic euxinic (anoxic and sulfidic) water masses prevailed in the mid-depth continental margins where sulfate content and primary productivity were relatively high, while the deep water remained persistently ferruginous5. However, recent studies have suggested a more anoxic and ferruginous subsurface ocean for the mid-Proterozoic (~ 1800–800 Ma). For instance, mass balance modeling of the redox-sensitive elements (RSEs) and the iron speciation data suggested the predominance of ferruginous water, rather than euxinia, in the shelf and slope environments9,11,12. Data of iron speciation and other redox proxies, such as cerium anomalies and uranium isotopes of carbonate rocks, as well as model calculations, further pointed out that in the very shallow water environments, even the wave-driven mixed portion of the ocean, the water mass was pervasively anoxic and ferruginous10,13,14,15. This pervasively anoxic and ferruginous subsurface ocean in the mid-Proterozoic was expected to cause extensive nitrate and phosphorus limitation, potentially inhibiting the ecological rise of eukaryotes14,16,17.
Despite the widespread anoxia, several studies have successfully tracked pulsed or pervasively oxic intermediate waters in the Mesoproterozoic ocean, making the ocean redox states of this time more enigmatic6,7,8,18,19. Distinct hypotheses have been put forward to interpret these mid-depth water oxygenation events, including fluctuating ocean oxygenations7,19, low oxygen consumption due to a weak biological pump6,20, and variations of ocean redox structures forced by climate dynamics18. The occurrences of oxygenated water mass in the generally anoxic mid-Proterozoic ocean may benefit the development of eukaryotes3. However, whether oxic/suboxic intermediate water masses occurred in the early Tonian ocean remains unclear12,14.
Here, to better understand the intricate ocean redox conditions and their regulatory mechanisms during the early Tonian Period, we present our comprehensive investigations on the marine siliciclastic and carbonate successions of the Huainan, Feishui, and Huaibei groups situated along the eastern margin of the North China craton (NCC). We propose that a shallow-water oxygen minimum zone may have occurred in the oligotrophic Tonian Xuhuai basin on the basis of sedimentological and petrological observations and multiple geochemical analyses. Such an ocean redox architecture could support the existence of a much larger bioavailable nitrate pool in the Xuhuai basin, potentially foreshadowing the ecological rise of eukaryotes in the middle and late Neoproterozoic if the redox architecture occurred in the early Tonian Xuhuai basin is confirmed to be a global phenomenon in future studies.
Results
Stratigraphy and sedimentological observations
As one of the oldest cratons in the world, the NCC contains unique and well-preserved Proterozoic geological records, with the Tonian sedimentary successions occurring extensively along its eastern margin21. Previous paleontological studies documented diverse eukaryotic fossils in these successions22,23,24, allowing us to investigate the intricate relationship between eukaryotic evolutions and ocean redox states. In this contribution, we focus on the Huainan and Feishui groups, which were deposited in the southern Xuhuai basin during the early Tonian Period (Fig. S1)23.
The Huainan Group, which unconformably overlies the Paleoproterozoic metamorphic basement of the Fengyang Group, consists of the Caodian, Bagongshan, and Liulaobei formations in ascending order (Fig. 1A). The Caodian and Bagongshan formations are mainly composed of conglomerates and sandstones, indicating deposition in fluvial to shoreface environments. The overlying Liulaobei Formation is lithologically subdivided into three intervals. While argillaceous and silty limestones characterize the lower and upper intervals, the middle interval of the Liulaobei Formation is predominately composed of finely laminated mudstones with thin calcareous mudstone interbeds (Fig. S2A–E). The common occurrences of cross-bedding in the lower and upper limestone intervals of the Liulaobei Formation (Fig. S2E) suggest deposition in a high-energy, shallow-water environment. In contrast, horizontal beddings and fine laminations in the shale and thin calcareous mudstone interbeds of the middle interval indicate deposition in a low-energy environment. The presence of slumping structures (Fig. S2C) and the absence of cross-bedding further suggest that the shales of the middle Liulaobei Formation were likely deposited in a mid-depth slope setting, when the sea level of the Xuhuai basin reached its highest extent during the early Tonian (Fig. 1A).

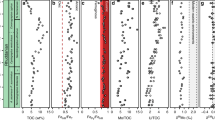
A Lithostratigraphy, biostratigraphy, sedimentary structures, geochronological data, and the proposed sea level changes. Note the proposed highest sea level occurs in the middle part of Liulaobei Formation. B Carbonate and organic matter carbon isotope profiles. Gray and black dots represent δ13Corg and δ13Ccarb, respectively, black dash line is the Lowess transgression of δ13Ccarb data fit with 90% confidence interval (blue shadow) calculated from standard errors. C, D Iron speciation profiles. Dashed lines in the FeHR/FeT profile represent calibrated thresholds indicative of oxic (<0.15) and anoxic (>0.38) water column conditions, and the dashed line in the FePy/FeHR profile denotes ferruginous water column condition (<0.6). E Ce anomaly profile. Dashed line represents the threshold (0.8) for distinguishing anoxic from oxic water column conditions. F Enrichment factors (EFs) for Mo, U, V, and Mn. The dashed line denotes where the EF value is one. G Baexcess contents (black dots) and Ba/Al ratios (gray dots). E—Ediacaran, Fm—Formation, BGS—Bagongshan, SX—Shouxian, JLQ—Jiuliqiao, FT—Fengtai. Radiometric dates sources: 1—ref. 28; 2—ref. 27; 3—ref. 26; 4—ref. 25. Source data are provided as a Source Data file.
The succeeding Feishui Group includes the Shouxian, Jiuliqiao, and Sidingshan formations in ascending order (Fig. 1A). The Shouxian Formation is dominated by calcareous quartz sandstones (Fig. S3A, I). The overlying Jiuliqiao Formation is distinguished by thin-bedded silty limestones (Fig. S3J) and is characterized by the common occurrences of intraclast (Fig. S3B), cross-bedding (Fig. S3C), and stromatolite reef (Fig. S3E). The Sidingshan Formation is mainly composed of dolostones, with common occurrences of stromatolite (Fig. S3F) and breccia (Fig. S3G), and is disconformably overlain by the Ediacaran Fengtai Formation diamictites (Fig. S3H). The Jiuliqiao Formation limestones and the Sidingshan Formation dolostones represent the deposition of shallow-water carbonate platform facies.
Bio- and chemostratigraphic data, as well as radiometric ages of sedimentary rocks, have robustly constrained the Huainan and Feishui groups to be early Tonian deposits21. A previous study reported a Sm-Nd isochron age of 801 ± 46 Ma for diagenetic cherts in the Sidingshan Formation25, providing a minimum depositional age for these successions. The presences of Trachyhystrichosphaera aimika and Sinosabellidites in the Liulaobei Formation and Pararenicola and Protoarenicola in the Jiuliqiao Formation indicate a Tonian age for these strata22,23,24, which is consistent with the U-Pb ages of the youngest detrital zircons obtained from the Bagongshan, Liulaobei, and Shouxian formations (Fig. 1)26,27,28. Additionally, carbonate carbon isotopic data (δ13Ccarb) of the Huainan and Feishui groups in the present study exhibit a pronounced negative excursion (up to ~9‰) in the middle part of the Liulaobei Formation (Fig. 1B). This negative excursion is comparable to the Majiatun anomaly observed in other regions of the NCC29,30,31, which has been constrained to occur between ~ 950 Ma and ~ 920 Ma by zircon ages of siliciclastics and intruded dolerite sills (Figs. S5 and S6)31. In addition, previously reported 87Sr/86Sr data for the carbonate rocks of the Huainan and Feishui groups are generally higher than 0.706032. These elevated values are comparable to those of the upper Huaibei and Jinxian groups, which have been suggested to record the highest peak of the seawater 87Sr/86Sr curve during the early Tonian (Figs. S5 and S6)33. Therefore, in conjunction with the above radiometric ages and paleontological data, the age of the Huainan Group can be approximately constrained from ~1000 Ma to ~920 Ma (see details in Supplementary Information).
For comprehensively understanding the oceanic chemistry of the Tonian Xuhuai basin, a total of 334 samples of the Huainan and Feishui groups were collected from three different sections in the Huainan region of the Xuhuai basin, including one drill core section (Shouxian drill core) which contains complete and continuous sedimentary records from the upper Bagongshan Formation to the lower Jiuliqiao Formation, and two outcrop sections (Baiguashan and Fengtai sections) that cover the remaining parts of the Feishui Group (Fig. S1). Eighty-one carbonate rock samples were also collected from the Shijia and Wangshan formations of the upper Huaibei Group (carbonate-dominated successions deposited in the Huaibei region of the Xuhuai basin) at the Shijiazhuang section (Fig. S1) to fill in the omission of previous chemostratigraphic studies29,33. Carbon isotope, major and trace element contents, and iron speciation analyses were conducted for carbonates and mudstones from the Liulaobei, Jiuliqiao, Sidingshan, Shijia, and Wangshan formations. All raw geochemical data both generated and used in the present study are provided in the Source Data file.
Geochemical data of samples from the Huainan and Feishui groups
The δ13Ccarb and δ13Corg (organic carbon isotopes) values and TOC (total organic carbon) contents of the analyzed samples range from −11.6‰ to 3.46‰, −31.28‰ to −24.82‰, and 0.02 wt.% to 0.23 wt.%, respectively. δ13Ccarb values exhibit a pronounced negative excursion (~9‰) in the middle interval of the Liulaobei Formation (Fig. 1B), accompanied by a less profound negative excursion of δ13Corg (~6‰, Fig. 1B) and a slight increase in TOC contents (~0.1 wt.%, Fig. S7) (see Supplementary Information for a detailed evaluation of diagenetic alterations).
Iron speciation measurements were conducted on 173 samples with FeT (total Fe contents) higher than 0.5 wt.% (Figs. 1 and S7). While FePy/FeHR ratios (FePy refers to pyrite Fe; FeHR refers to highly reactive Fe) are generally below 0.6, FeHR/FeT ratios display significant variations (Fig. 1C, D). In the lower interval of the Liulaobei Formation, FeHR/FeT values range from 0.09 to 0.42 (n = 14), with the majority falling between 0.15 and 0.38 (n = 10). The middle interval of the Liulaobei Formation yields persistently low FeHR/FeT values, generally below the threshold value of 0.15 (53 out of 79 samples). FeHR/FeT values are more variable throughout the upper interval of the Liulaobei Formation, ranging from 0.04 to 0.76 (n = 40). The Jiuliqiao and lower Sidingshan formations exhibit consistently high FeHR/FeT ratios that exceed the threshold value of 0.38, with an average of 0.57 (n = 41).
Rare earth elements and yttrium (REY) concentrations were analyzed for 53 carbonate rock samples from the Liulaobei, Jiuliqiao, and Sidingshan formations, using the leaching method established by Tostevin et al.34. High Y/Ho ratios and low total REY contents (ΣREY) are identified for all samples (Fig. S8). The Eu anomaly values [Eu/Eu* = 2EuN/(SmN + GdN), XN refers to the PAAS-normalized REY abundance] vary from 1.11 to 17.28 and exhibit strong correlations with the Ba/Nd values (R2 = 0.994, Fig. S9), indicating that these positive Eu anomalies are actually caused by the significant interference effect of high barium content during instrumental analysis rather than by hydrothermal alterations35. The fractionation between light rare earth elements (LREEs) and heavy rare earth elements (HREEs) varies among different formations (Fig. S10). The REY distribution patterns for samples from the Liulaobei Formation exhibit LREEs depletion or nearly flat shape, with PrN/ErN ratios ranging from 0.37 to 1.03 (0.62 on average, n = 23), while samples of the Jiuliqiao Formation exhibit both LREEs enrichment and depletion, with PrN/ErN ratios ranging from 0.47 to 1.52 (0.87 on average, n = 17). The Sidingshan Formation dolostones show the most significant LREEs depletion, with PrN/ErN ratios ranging from 0.30 to 0.90 (0.48 on average, n = 13). The Ce anomaly is calculated as Ce/Ce* = CeN/PrN2/NdN, following the ref. 36, and the Ce/Ce* values for all analyzed samples range from 0.72 to 1.43, with an average of 1.15 (n = 53) (Fig. 1E).
RSEs measurements were carried out for fine-grained siliciclastic samples from the middle interval of the Liulaobei Formation. The concentrations of U, Mo, V, and Mn range from 1.63 to 2.58 ppm (n = 45), 0.15 to 1.57 ppm (n = 45), 61.34 to 124.03 ppm (n = 99), and 74.80 to 10,626.15 ppm (n = 99), respectively. Enrichment factors [EF(X) = (Xsample/Alsample)/(Xaverage shale/Alaverage shale)] were used to standardize RSEs’ concentrations37. Nearly all EFs of U, Mo, and V are less than one, except for two samples that exhibit EF(Mo) of 1.3 and EF(V) of 1.0, respectively, while the EFs of Mn range from 0.1 to 17.0 and average at 1.2 (n = 99) (Fig. 1F).
The redox states of the depositional environment for Huainan and Feishui groups
As a well-calibrated technique, iron speciation ratios have been successfully employed to determine the local basin redox conditions from which the fine-grained siliciclastic rocks are deposited38, and recent studies have demonstrated their applicability to carbonate rocks containing sufficient total iron (FeT >0.5 wt.%)39. Some acknowledged caveats exist in the application of iron-based redox proxy. Rapid sedimentation, common during turbidite deposition, could obscure the primary iron enrichments and result in an oxic illusion40, indicating that the utilization of iron speciation analyses is best within the context of sedimentological conditions41. The conversion of FeHR to authigenic iron-rich clays during digenetic or metamorphic processes is another case that could diminish the primary depositional FeHR/FeT ratio42. Such circumstances can be avoided by utilizing a reduced threshold value of 0.15 for FeHR/FeT43. Fe/Al ratios can also be used to assess this process since this ratio is not affected by the redistribution of iron during diagenesis41. In this study, FeHR/FeT < 0.15 and FeHR/FeT > 0.38 indicate oxic and anoxic conditions, respectively43,44. In the latter case, FePy/FeHR > 0.8 reflects euxinic settings, whereas FePy/FeHR < 0.6 indicates sediments deposited under ferruginous water columns41,45.
The iron speciation data of the Huainan and Feishui groups show systematic variations throughout the succession. Particularly noteworthy is the predominately oxic setting assigned for the deposition of the middle Liulaobei Formation mudstones (FeHR/FeT ratios average 0.13 ± 0.06, Fig. 1C), which contrasts with the prevalently anoxic subsurface ocean proposed for the Xuhuai basin in previous studies12,13,14. Iron speciation data for samples from the carbonate intervals of the lower and upper Liulaobei Formation, the entire Jiuliqiao Formation, and the lower part of the Sidingshan Formation generally show no evidence for oxic seawater, with most FeHR/FeT ratios falling into the equivocal and anoxic zones (0.49 ± 0.13, Fig. 1C). Slight increases of the highly reactive iron contents in carbonate, iron oxide, and magnetite are also found in these carbonate rock samples relative to the mudstones (Fig. S7). Consistently low FePy/FeHR ratios (0.12 ± 0.13) imply non-euxinic conditions during carbonate deposition (Fig. 1D).
Most studied samples with FeHR/FeT ratio less than 0.15 display Fe/Al ratios within the range that is indicative of oxic depositional settings (0.44–0.66)39, suggesting the absence of authigenic iron enrichment in silicate minerals (Fig. S7). However, some samples with equivocal (0.15–0.38) and anoxic (>0.38) FeHR/FeT signals from the lower and upper intervals of the Liulaobei Formation also exhibit Fe/Al ratios falling into the oxic zone, and samples from the Jiuliqiao and lower Sidingshan formations, with iron speciation data showing anoxic setting, yield Fe/Al ratios far below 0.44 (Fig. S7). This phenomenon can be attributed to the low background Fe/Al ratio in the parent rocks41 or the removal of iron during diagenetic processes. It should be noted that some other studies on Neoproterozoic sediments have also reported anomalously low Fe/Al ratios46,47,48, and their origin remains to be investigated. Indeed, in the modern ocean with oxygen minimum zones (OMZs), e.g., the Peruvian continental margin, sediments deposited below the lower boundary of the OMZ generally exhibit higher Fe/Al ratios compared to those within the OMZ due to the net supply of iron from the OMZ to the slope area49.
The REY pattern can also function as an independent redox indicator for carbonate rocks due to the distinctive redox behaviors of Ce relative to its neighboring rare earth elements (REEs) in the water column34. Under oxic conditions, Ce can be easily scavenged from the water, resulting in a negative Ce anomaly in seawater (Ce/Ce* < 0.8). While under anoxic settings, the Ce anomaly is either absent or weak. In the present study, the generally high Y/Ho ratios, low ΣREY, and low LREEs/HREEs ratios (Figs. S8 and S10) suggest successful extractions of the original seawater signal in our experiments34. The lack of correlations between detritus proxies (e.g., Al and Th contents) and Y/Ho ratios, ΣREY, and Ce/Ce* values further exclude the possible contamination from partial dissolutions of terrigenous detritus (Fig. S11)34. The persistent absence of negative Ce anomalies (1.15 ± 0.11), aligning with the iron speciation data, suggests an anoxic depositional setting for the carbonate rocks of Jiuliqiao and Sidingshan formations. The absence of negative Ce anomalies for carbonate rocks from the lower and upper intervals of the Liulaobei Formation also confirms the predominately anoxic states deduced from their FeHR/FeT values between 0.15 and 0.38 (Fig. 1).
RSEs provide additional helpful insights into the redox conditions of the water column where siliciclastic rocks were deposited. Conventionally, the consistently low EFs of Mo (0.37 ± 0.24), V (0.78 ± 0.09), and U (0.68 ± 0.08) in the middle Liulaobei Formation may indicate the presence of persistently oxic settings (Fig. 1F)37. The depletion of V, which is only found in modern sediments under oxygenated water columns, is a robust indicator of bottom water oxygenation7. Consistent with the sediments deposited beneath suboxic bottom water in the modern eastern tropical Pacific, the middle Liulaobei Formation mudstones also exhibit a general enrichment of U relative to Mo (Figs. 1 and S12)50. Furthermore, the retention of Mn in sediments typically requires well-oxygenated conditions51,52. Thus, mild enrichments of Mn (1.20 ± 2.35, Fig. 1F) in the middle interval of the Liulaobei Formation may also indicate oxic bottom water conditions51,52. It is worth noting that some studies suggested that widespread anoxia in the mid-Proterozoic ocean would have drawn down the concentrations of the redox-sensitive elements, potentially limiting the use of RSEs, particularly U and Mo, in determining local redox states11,53. Notwithstanding, previous works have successfully discriminated different enrichment degrees of RSEs in the Proterozoic anoxic and oxic bottom waters18,54. In the present study, while the U and Mo concentrations of the middle Liulaobei Formation mudstones are generally depleted relative to the average shales, the Shijia and Wangshan formations, which were deposited contemporaneously with the middle Liulaobei Formation but beneath anoxic water columns (see details below), exhibit fluctuant EFs values, with many exceeding one, further affirming the usefulness of RSEs in reconstructing paleo-redox conditions in the Precambrian period. Taken together, the RSEs data for the shales/silty mudstones of the Liulaobei Formation are generally consistent with the iron speciation data, indicating the presence of oxic/suboxic conditions for the deposition of the middle Liulaobei Formation.
Different from this study, Guilbaud et al.12 demonstrated a pervasively ferruginous condition for the deposition of the Liulaobei Formation based on iron speciation data12. Our data show that the middle interval of the Liulaobei Formation, which was deposited under oxic bottom water conditions as indicated by the abovementioned redox proxies, also exhibits negative δ13Ccarb (~9‰) and δ13Corg (~6‰) excursions (Fig. 1B). The coupled carbon isotope negative excursions are absent in the Liulaobei Formation profiles reported by Guilbaud et al.12,17, suggesting that the shale/silty mudstones-dominated middle Liulaobei Formation may have been omitted in their study. Their studied samples (from the stratigraphic interval of ~170 m out of ~470 m of the Liulaobei Formation and generally with low Al contents12,17) may only represent the upper carbonate interval of the Liulaobei Formation in the present study.
Spatial heterogeneity of the redox states in the Xuhuai basin
The iron speciation, RSEs, and REY data indicate substantial variations in marine redox states during the deposition of the Huainan and Feishui groups (Fig. 1). These variations can be empirically interpreted as reflections of secular redox evolutions of the Xuhuai basin. Nonetheless, the variation of our redox proxy data also correlates strongly with depositional water depth. That is to say, shales and silty mudstones in the middle Liulaobei Formation were deposited in oxic mid-depth slope environments, while carbonate rocks in the lower and upper intervals of the Liulaobei Formation, as well as the Jiuliqiao and lower Sidingshan formations, were deposited in persistently anoxic and ferruginous shallow-water settings (Figs. 1 and 2). Whether this relationship reflects inherent redox heterogeneities among different environments or merely the consequences of periodic water column oxygenations requires basin-scale investigations.

A Crossplot of FePy/FeHR versus FeHR/FeT. Note that most mudstones (purple dots) of the Liulaobei Formation fall into the oxic zone, whilst the majority of shallow-water carbonate rocks (blue triangles and black diamonds) from the Huainan, Feishui, and Huaibei groups fall into the ferruginous domain. B Plot of FeHR/FeT versus Al contents. The significant negative correlation may suggest transformation from anoxic to oxic settings with depth increase. Dashed lines indicate the boundaries for distinguishing different water column redox conditions. Iron speciation data of the carbonate rocks from the Huaibei region are from ref. 14 (solid black diamonds) and this study (hollow black diamonds). Source data are provided as a Source Data file.
Here, we use the Huaibei Group distributed in the Huaibei region to assess the spatial heterogeneity of redox states in the Xuhuai basin (Fig. S1B). Previous studies suggested that the deposition of the Huaibei Group is approximately contemporaneous with the Huainan and Feishui groups29. The δ13Ccarb data reported in the present study demonstrate the occurrences of the Majiatun anomaly (~950–920 Ma), a remarkable carbon isotope negative excursion that occurs in the early Tonian successions31, in both the Shijia Formation of the upper Huaibei Group and the Liulaobei Formation of the Huainan Group, providing direct evidence for a more precise stratigraphic correlation between them (Fig. S5). This stratigraphic correlation is further supported by lithostratigraphic and 87Sr/86Sr chemostratigraphic data of the Huaibei, Huainan, and Feishui groups (see Supplementary Information for detailed discussions on stratigraphic correlations).
Unlike the Huainan and Feishui groups that encompass both shallow- and mid-depth-water deposits23, the Huaibei Group was exclusively deposited in the shallow-water environment of the Xuhuai basin29. Our iron speciation data of the Shijia and Wangshan formations, along with previously reported iron speciation and nitrogen isotope data14, suggest a persistently anoxic and ferruginous shallow-water condition during the deposition of the entire Huaibei Group (Figs. 2 and S13). This long-lasting ferruginous condition accords with the depositional environment of the shallow-water carbonates in the Huainan and Feishui groups (Fig. 2), further indicating the spatial heterogeneity of marine redox states in the Xuhuai basin during the deposition of the middle interval of the Liulaobei Formation.
Unfortunately, the absence of deposition in mid-depth environments before and after the deposition of the Liulaobei Formation hinders further investigations of the basinal redox structures in the Xuhuai basin. Given the generally anoxic condition assigned to the early Neoproterozoic subsurface ocean in previous studies13,14, the occurrence of the oxic/suboxic mid-depth water masses in the Xuhuai basin may reflect a transient ocean oxygenation event. However, ocean oxygenation generally commences with increases in oxygen levels in surface water and gradually extends from shallow to deep waters55. Substantial geochemical changes would be expected when an oxygenation event happened, which is inconsistent with the absence of distinct variations in δ238U and δ15N values in the Huaibei Group13,14. This phenomenon may indicate that the oxic/suboxic mid-depth water masses recorded in the middle Liulaobei Formation mudstones were not formed during pulsed oceanic oxygenation events but more likely represent an inherent oceanic redox characteristic of the early Tonian Xuhuai basin. The variation of the redox proxy data of the Huainan and Feishui groups thus plausibly reflects the spatial heterogeneity of marine redox conditions within the Xuhuai basin, rather than the secular redox evolutions of the depositional basin.
It is noteworthy that no direct evidence for oxic shallow water was obtained in the present study, although it has been suggested that the surface ocean became oxygenated after the Great Oxidation Event5. This phenomenon could be attributed to the persistent existence of anoxic and ferruginous zones beneath the surface oxic layer in shallow-water environments. Though no Ce negative anomalies (i.e., no oxic conditions) are identified in the shallow-water carbonate rock samples, the existence of the oxic surface water in the Xuhuai basin can be partly inferred from the slight LREEs depletion preserved in the shallowest Sidingshan Formation dolostones compared to the nearly flat or even right deviation REE pattern in the relatively deeper Jiuliqiao Formation limestones (Fig. S10). The broad LREEs depletions in the carbonate rocks of the Liulaobei Formation could be ascribed to the proximity of the depositional location to mid-depth oxic/suboxic water masses with increasing depth (Figs. S8 and S10). Given the deep water was anoxic in the mid-Proterozoic ocean5, multiple redox proxies can be used to conceptualize a more complex oxic upper ocean in the Xuhuai basin, where the persistently shallow anoxic and ferruginous water wedge was sandwiched between the oxic/suboxic mid-depth water masses and the surface mixed layer (Fig. 3).

A Idealized basin topography and Proterozoic stratified ocean. Blue dashed line marks the chemocline between the oxic upper ocean and the anoxic deep ocean. Khaki shaded area represents the basement rock. B Schematic representation of the shallow-water oxygen minimum zone (OMZ, green shaded area) in the Xuhuai basin. The relatively higher primary productivity in the nearshore area consumed the dissolved oxygen and maintained the prevalent shallow-water OMZ in an oligotrophic ocean. The insignificant riverine sulfate input relative to the FeHR during this period further prohibited the microbial sulfate reduction and resulted in a ferruginous OMZ. The inset in B shows the proposed variations of O2 (solid black line), Fe2+ (dashed black line), and H2S (dotted gray line) in the water column marked by a dotted rectangle.
The oxygen level of ocean subsurface water is ultimately determined by the balance between O2 supply from the atmosphere and O2 consumption by aerobic respiration and other reduced species within the water column20. In some areas of the oxic modern ocean, where the surface production is elevated and the circulation of deep oxygen-rich water is sluggish, the quick consumption of the dissolved oxygen through aerobic degradation of organic matter could form steady-state oxygen-limited zones in the mid-depth water column, i.e., the OMZs56. The conceptual redox model for the Xuhuai basin during the early Tonian, or at least during the deposition of the middle Liulaobei Formation, may resemble the modern ocean’s OMZ redox architecture but developed at a much shallower location (Fig. 3).
Discussion
The Proterozoic atmospheric O2 concentrations are undoubtedly well below the present atmospheric level (PAL)5. While an increase in the atmospheric O2 level could certainly facilitate the formation of oxic mid-depth water masses, a single oxygenation event cannot fully explain the spatial heterogeneity of marine redox states in the Tonian Xuhuai basin. Indeed, model calculations speculate that even with low atmospheric O2 concentrations (e.g., 1% PAL), large seafloor regions could also maintain mildly oxygenated conditions in an oligotrophic ocean20. With low sulfide and ferrous iron contents, the mid-Proterozoic ocean was suggested to lack significant redox buffers8. In this context, lateral variations in primary productivity could account for the spatial heterogeneity of marine redox states. That is to say, negligible primary productivities in the offshore ocean could effectively suppress aerobic respiration in the water column, allowing sufficient oxygen diffusion and resulting in persistent oxic/suboxic intermediate water. Conversely, the slight flourish of microorganisms and their oxygen-consuming remineralizations on the carbonate platforms, e.g., in the Xuhuai basin, may ultimately lead to the formation of shallow-water OMZs.
Biogenic barium (Babio) is a widely used paleo-productivity proxy in deep-time oceanographic studies57. In the ocean, the flux of particulate barium is closely linked to that of particulate organic matter58, which is attributed to the localized development of microenvironments that promote the formation of barite during organic matter decay59. Unlike other paleo-productivity proxies (e.g., TOC and organic phosphorus), Babio is not sensitive to sediment bulk accumulation rates (BAR)57, making it a good paleo-productivity indicator for sedimentary successions without precise age constraints. In practice, the Babio is often substituted by excess barium (Baexcess) and Ba/Al ratios. Providing that all non-detrital Ba is biogenic, the Baexcess value is determined by the following equation: Baexcess = BaT–AlT*(Ba/Al)Det. BaT and AlT refer to the total Ba and Al concentrations in each sample, and (Ba/Al)Det is for average sediments, which is assumed to be 0.003760.
Systematic deviations of Baexcess and Ba/Al ratios were observed between carbonate rocks and mudstones in the Huainan and Feishui groups. Mudstones of the middle Liulaobei Formation deposited in oxic mid-depth environments typically yield near zero or even negative Baexcess values (−0.84 ± 31.37, n = 47) and extremely low Ba/Al ratios (0.0037 ± 0.0004, n = 47), while Baexcess values (157.83 ± 158.52, n = 64) and Ba/Al ratios (0.0089 ± 0.0061, n = 64) of carbonate rocks deposited in anoxic shallow-water environments are generally high (Fig. 1G). Relatively higher Baexcess values (18.79 ± 43.81, n = 19) and Ba/Al ratios (0.0101 ± 0.0114, n = 19) are also identified in the Shijia and Wangshan formations, further indicating higher primary productivity in nearshore than in offshore areas of the Xuhuai basin during the deposition of the middle Liulaobei Formation. The slight increase in export production would rapidly deplete the dissolved oxygen in the water column, ultimately resulting in the formation of shallow-water OMZs in the carbonate platforms (Fig. 3). The low riverine sulfate input further inhibited microbial sulfate reduction, which restricted the shallow water anoxic zone to be ferruginous rather than euxinic12.
The spatial heterogeneity of primary productivity in the early Tonian Xuhuai basin might be attributed to a seaward decreasing nutrient gradient. Phosphorus is one of the critical macro-nutrients that ultimately regulates ocean primary productivity61, and its concentration in the oceans is controlled by marine redox states and continental weathering inputs62. The retention of phosphorus in sediments is enhanced under ferruginous water columns due to the increased uptake by iron minerals and the suppression of sulfate reduction in pore water17. As a result, the widespread ferruginous OMZ in the nearshore area of the Xuhuai basin can significantly influence the lateral distribution of phosphorus. While the constant weathering inputs could maintain a stable bioavailable phosphorus pool in the nearshore area, the persistent scavenging of phosphorus in the ferruginous shallow-water OMZs would prohibit the lateral transport of reactive phosphorus, resulting in a decrease in phosphorus accumulation and substantial limitations of primary productivity in the offshore area.
Notably, consistent with the anoxic shallow-water environment in the Xuhuai basin, previous studies have successfully revealed the occurrences of shallow marine anoxia and euxinia in other late Tonian basins (~800 Ma)63,64. Indeed, considering that the releases of phosphorus from sediments to the water column are particularly intense under euxinic conditions65, the presence of euxinic shallow-water settings may significantly promote the elevation of the primary productivity and the expansion of anoxic water masses in the ocean, which would eventually inhibit the development of oxic/suboxic mid-depth water masses. Consistent with this inference, a recent study of the Mesoproterozoic Xiamaling Formation in the North China craton indicated that while euxinic and ferruginous water masses alternately occurred in the Yanliao basin, oxic/suboxic mid-depth water masses only occurred under ferruginous OMZs18.
The Liulaobei and Jiuliqiao formations yield abundant macroscopic carbonaceous compression fossils22,24. Notably, in addition to the common and possibly planktonic Chuaria, Tawuia, and Sinosabellidites, the Jiuliqiao Formation also contains benthic sessile algal fossils, including Protoarenicola and Pararenicola, which possibly suggest the presence of free oxygen in the bottom water22,24. This inference seems inconsistent with the anoxic conditions for the deposition of the Jiuliqiao Formation, as discussed above. Considering that horizons containing in situ benthic algal fossils are limited and sporadic, a plausible scenario is that the distribution of the shallow-water OMZ in the Xuhuai basin is dynamic due to the fluctuating factors that control the primary production. During the transient contraction of the shallow-water OMZ, the inflow of oxic and probably nitrate-rich mid-depth water masses to the shallow photic zone would create habitable benthic niches. High-resolution redox proxy analyses focused on layers with benthic algal fossils are needed to test such changes.
The early Tonian witnessed the transition from a prokaryote-dominated to a eukaryote-dominated marine ecosystem1,2 and many pivotal evolutionary innovations of eukaryotes3. These processes are likely to be associated with changes in nutrient supplies and redox states. In the phosphorus-limited mid-Proterozoic ocean16,62, bioavailable nitrate was hypothesized to play a crucial role in regulating the evolution of eukaryotes14. Nitrogen cycles are deeply linked to marine redox states, and nitrate availability in the ocean is regarded as the key factor controlling the competition between eukaryotic and prokaryotic primary producers66. Therefore, while the global availability of nitrate and bioactive phosphorus may not have undergone substantial increases until ~800 Ma14,16, the occurrence of oxic/suboxic and generally oligotrophic mid-depth water column would have provided constant supplies of bioavailable nitrate67 and inhibited the denitrification and anammox68, ultimately resulting in the formation of a stable nitrate pool in the offshore area. The stable and higher δ15N values obtained from the Liulaobei Formation (4.29 ± 0.13‰)69 compared to those from the contemporaneous Huaibei Group (2.03 ± 0.87‰)14 further confirm this inference, as lateral variations in δ15N are generally indicative of spatial heterogeneity of nitrogen cycling and redox conditions in the ocean70. While high δ15N records can be successfully preserved in mudstones deposited beneath oxic/suboxic and nitrate-rich mid-depth water masses, slight increases of the primary productivity in the shallow-water OMZ would quantitatively remove nitrates in the local water column and thus lead to near-zero δ15N signals in shallow-water deposits. We emphasize that the proposed nitrate distribution pattern does not contradict the previous study, which suggested a decline in nitrate contents with increasing water-depth70, but virtually reflects a more complex δ15N distribution pattern in an oxic/suboxic upper ocean.
As discussed above, the sandwiched marine redox structure plausibly represents an inherent characteristic of the Xuhuai basin. However, a possible oxygenation event, which would undoubtedly benefit the formation of the oxic mid-depth water, cannot be completely excluded during the deposition of the middle Liulaobei Formation. Future work is still needed to test the duration of the proposed redox structure in the Xuhuai basin and its potential global distribution. If the shallow-water oxygen minimum zone that occurred in the early Tonian Xuhuai basin is confirmed to be a global phenomenon in the future, we suggest that such a marine redox landscape could create essential ecological niches for eukaryotes, foreshadowing their ultimately ecological rise in the middle and late Neoproterozoic.
Methods
TOC and C isotopes
Total organic carbon (TOC) contents and organic carbon isotopic analyses were performed with Delta V Advantage mass spectrometer. Before analyses, the sample was first decarbonated with sufficient 3% M HCl for 24 h, then thoroughly rinsed three times with ultrapure water and oven-dried. The TOC content and δ13Corg analytical precisions are better than 5% and 0.2‰ (1σ), respectively. Carbonate carbon and oxygen isotopic analysis was conducted with a MAT 253 isotope ratio mass spectrometer and the analytical precision is better than 0.04‰ (1σ). The carbon and oxygen isotopic compositions of the calcareous mudstones from the middle Liulaobei Formaion were analyzed at the State Key Laboratory of Oil and Gas Reservoir Geology and Exploitation (Chengdu University of Technology) with the following process. Approximately 0.5 mg of rock powder was treated with 0.25 ml of pure phosphoric acid at 40 °C for 4 h. The resulting CO2 gas was separated and collected by an Iso Flow instrument, and directly analyzed in an Elementar Isoprime precisION mass spectrometer. Analytical precisions for carbon and oxygen isotopes are better than 0.2‰.
Rare earth elements
To avoid the potential contamination from non-carbonate phases, Rare earth elements and yttrium were extracted using 2% (w/v) nitric acid following the sequential dissolution method developed by ref. 34. The sample powder containing ~60 mg carbonate was first cleaned with ultrapure water and then treated with sufficient nitric acid to digest ~20% carbonate. Whereafter, the sample was put into the shaker for 20 min at room temperature before being centrifuged. The residue was rinsed three times with ultrapure water to remove adsorbed ions. Then, the residue was treated with sufficient nitric acid again to digest the following ~40% carbonate. After agitating on the shaker for 20 min, the supernatant was collected and analyzed with inductively coupled plasma mass spectrometry (ICP-MS, Agilent 7700x). The analytical precision is better than 5%. The concentration of the rare earth elements in carbonate components was determined by dividing their masses in the leachate by the corresponding weight of carbonate, which can be calculated based on the Ca and Mg contents of the same leachate.
Iron speciation
Iron content in pyrite (Fepy) was stoichiometrically calculated from the weight percentage of Ag2S, which was extracted using the chromium reduction method. Iron in carbonate, magnetite, and oxides were extracted from ~0.1 g powdered sample within 15 ml centrifuge tubes following the well-established iron sequential extraction schemes38. The sample powder was treated with a sodium acetate solution at 50 °C (pH 4.5 for 48 h) to obtain carbonate-associated Fe (Fecarb); Then, the residue was treated with a sodium dithionite solution at room temperature (pH 4.8 for 2 h) to get Fe (oxyhydr)oxide minerals (Feox); At last, the residue was treated with ammonium oxalate (at room temperature for 6 h) to get magnetite (Femag). The supernatant in each extraction was removed through a centrifugation step. Iron concentrations of these extracted species were measured using an inductively coupled plasma optical emission spectrometer (ICP-OES, Agilent 710). The analytical precision is better than 5%.
Major and trace elements
Major elements of the whole-rock samples were analyzed at the ALS Chemex (Guangzhou) Co., Ltd and the Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences (NIGPAS) with the following two methods, respectively. (1) Analysis on fused lithium borate glass disk using X-ray fluorescence spectroscopy (XRF, PANalytical PW2424) was carried out for carbonates, and (2) concentrated HF–HNO3 digestion followed by analysis using ICP-OES (Agilent 710) was performed for the mudstones. The analytical precision is better than 5%.
Trace element concentration analysis for the mudstone was carried out at the Wuhan SampleSolution Analytical Technology Co., Ltd. The sample was first completely digested by concentrated HF and HNO3 and measured by ICP-MS (Agilent 7700e). The measurement precision is better than 5%.
Unless otherwise indicated, sample preparations and analyses were carried out at NIGPAS.
Data availability
Source data are provided with this paper.
References
Knoll, A. H., Javaux, E. J., Hewitt, D. & Cohen, P. Eukaryotic organisms in Proterozoic oceans. Philos. Trans. R. Soc. B 361, 1023–1038 (2006).
Porter, S. M. Insights into eukaryogenesis from the fossil record. Interface Focus 10, 20190105 (2020).
Xiao, S. & Tang, Q. After the boring billion and before the freezing millions: evolutionary patterns and innovations in the Tonian Period. Emerg. Top. Life Sci. 2, 161–171 (2018).
Cohen, P. A. & Kodner, R. B. The earliest history of eukaryotic life: uncovering an evolutionary story through the integration of biological and geological data. Trends Ecol. Evol. 37, 246–256 (2022).
Lyons, T. W., Reinhard, C. T. & Planavsky, N. J. The rise of oxygen in Earth’s early ocean and atmosphere. Nature 506, 307–315 (2014).
Sperling, E. A. et al. Redox heterogeneity of subsurface waters in the Mesoproterozoic ocean. Geobiology 12, 373–386 (2014).
Zhang, S. et al. Sufficient oxygen for animal respiration 1400 million years ago. Proc. Natl Acad. Sci. USA 113, 1731–1736 (2016).
Planavsky, N. J. et al. Evidence for episodic oxygenation in a weakly redox-buffered deep mid-Proterozoic ocean. Chem. Geol. 483, 581–594 (2018).
Gilleaudeau, G. J. et al. Uranium isotope evidence for limited euxinia in mid-Proterozoic oceans. Earth Planet. Sci. Lett. 521, 150–157 (2019).
Liu, X. et al. A persistently low level of atmospheric oxygen in Earth’s middle age. Nat. Commun. 12, 351 (2021).
Reinhard, C. T. et al. Proterozoic ocean redox and biogeochemical stasis. Proc. Natl Acad. Sci. USA 110, 5357–5362 (2013).
Guilbaud, R., Poulton, S. W., Butterfield, N. J., Zhu, M. & Shields-Zhou, G. A. A global transition to ferruginous conditions in the early Neoproterozoic oceans. Nat. Geosci. 8, 466–470 (2015).
Zhang, F. et al. Uranium isotope evidence for extensive shallow water anoxia in the early Tonian oceans. Earth Planet. Sci. Lett. 583, 117437 (2022).
Kang, J., Gill, B., Reid, R., Zhang, F. & Xiao, S. Nitrate limitation in early Neoproterozoic oceans delayed the ecological rise of eukaryotes. Sci. Adv. 9, eade9647 (2023).
Reinhard, C. T., Planavsky, N. J., Olson, S. L., Lyons, T. W. & Erwin, D. H. Earth’s oxygen cycle and the evolution of animal life. Proc. Natl Acad. Sci. USA 113, 8933–8938 (2016).
Planavsky, N. J. et al. A sedimentary record of the evolution of the global marine phosphorus cycle. Geobiology 21, 168–174 (2023).
Guilbaud, R. et al. Phosphorus-limited conditions in the early Neoproterozoic ocean maintained low levels of atmospheric oxygen. Nat. Geosci. 13, 296–301 (2020).
Song, Y. et al. Dynamic redox and nutrient cycling response to climate forcing in the Mesoproterozoic ocean. Nat. Commun. 14, 6640 (2023).
Yang, X. et al. Fluctuating oxygenation and dynamic iron cycling in the late Paleoproterozoic ocean. Earth Planet. Sci. Lett. 626, 118554 (2024).
Reinhard, C. T. & Planavsky, N. J. The history of ocean oxygenation. Annu. Rev. Mar. Sci. 14, 331–353 (2022).
Pang, K. et al. Integrated meso-neoproterozoic stratigraphy in the Jiao-Liao-Xu-Huai area of North China Craton: a review. J. Stratigr. 45, 467–492 (2021).
Dong, L. et al. Restudy of the worm-like carbonaceous compression fossils Protoarenicola, Pararenicola, and Sinosabellidites from early Neoproterozoic successions in North China. Palaeogeogr. Palaeoclimatol. Palaeoecol. 258, 138–161 (2008).
Tang, Q. et al. Organic-walled microfossils from the early Neoproterozoic Liulaobei Formation in the Huainan region of North China and their biostratigraphic significance. Precambrian Res. 236, 157–181 (2013).
Li, G. et al. Tonian discoidal fossils from North China: relating discs to worm-like annulated tubes and their paleoecological and evolutionary implications. Palaeogeogr. Palaeoclimatol. Palaeoecol. 628, 111780 (2023).
Yang, J., Zheng, W., Tao, X. & Wang, Z. The Sm-Nd age of cherts from Sidingshan Formation of the Huainan Group of Anhui Province. Geol. Rev. 50, 413–417 (2004).
Zhao, H. et al. New geochronologic and paleomagnetic results from early Neoproterozoic mafic sills and late Mesoproterozoic to early Neoproterozoic successions in the eastern North China Craton, and implications for the reconstruction of Rodinia. Geol. Soc. Am. Bull. 132, 739–766 (2020).
Li, G. et al. The characteristics of LA-ICP-MS detrital zircon U-Pb age from the Meso-Neoproterozoic strata in Huainan area and their geological significance. J. Stratigr. 45, 115–141 (2021).
Sun, F. et al. Provenance analysis of the late Mesoproterozoic to Neoproterozoic Xuhuai Basin in the southeast North China Craton: implications for paleogeographic reconstruction. Precambrian Res. 337, 105554 (2020).
Xiao, S. et al. Biostratigraphic and chemostratigraphic constraints on the age of early Neoproterozoic carbonate successions in North China. Precambrian Res. 246, 208–225 (2014).
Park, H. et al. Deposition age of the Sangwon Supergroup in the Pyongnam basin (Korea) and the Early Tonian negative carbon isotope interval. Acta Petrol. Sin. 32, 2181–2195 (2016).
Zhang, Z. et al. Oldest-known Neoproterozoic carbon isotope excursion: earlier onset of Neoproterozoic carbon cycle volatility. Gondwana Res. 94, 1–11 (2021).
Yang, J., Zheng, W., Wang, Z. & Tao, X. Age determining of the upper Precambrian System of northern Jiangsu–Anhui by using Sr and C isotopes. J. Stratigr. 25, 44–47 (2001).
Zhou, Y. et al. Reconstructing Tonian seawater 87Sr/86Sr using calcite microspar. Geology 48, 462–467 (2020).
Tostevin, R. et al. Effective use of cerium anomalies as a redox proxy in carbonate-dominated marine settings. Chem. Geol. 438, 146–162 (2016).
Jarvis, K. E., Gray, A. L. & McCurdy, E. Avoidance of spectral interference on europium in inductively coupled plasma mass spectrometry by sensitive measurement of the doubly charged ion. J. Anal. Spectrom. 4, 743–747 (1989).
Lawrence, M. G., Greig, A., Collerson, K. D. & Kamber, B. S. Rare earth element and yttrium variability in South East Queensland waterways. Aquat. Geochem. 12, 39–72 (2006).
Tribovillard, N., Algeo, T. J., Lyons, T. & Riboulleau, A. Trace metals as paleoredox and paleoproductivity proxies: an update. Chem. Geol. 232, 12–32 (2006).
Poulton, S. W. & Canfield, D. E. Development of a sequential extraction procedure for iron: implications for iron partitioning in continentally derived particulates. Chem. Geol. 214, 209–221 (2005).
Clarkson, M. O., Poulton, S. W., Guilbaud, R. & Wood, R. A. Assessing the utility of Fe/Al and Fe-speciation to record water column redox conditions in carbonate-rich sediments. Chem. Geol. 382, 111–122 (2014).
Raiswell, R. & Canfield, D. E. Sources of iron for pyrite formation in marine sediments. Am. J. Sci. 298, 219–245 (1998).
Poulton, S. W. The Iron Speciation Paleoredox Proxy (Cambridge University Press, 2021).
Lyons, T. W. & Severmann, S. A critical look at iron paleoredox proxies: new insights from modern euxinic marine basins. Geochim. Cosmochim. Acta 70, 5698–5722 (2006).
Raiswell, R. et al. Turbidite depositional influences on the diagenesis of Beecher’s Trilobite Bed and the Hunsrück Slate; sites of soft tissue pyritization. Am. J. Sci. 308, 105–129 (2008).
Poulton, S. W. & Canfield, D. E. Ferruginous conditions: a dominant feature of the ocean through Earth’s history. Elements 7, 107–112 (2011).
Benkovitz, A. et al. Tracing water column euxinia in Eastern Mediterranean Sapropels S5 and S7. Chem. Geol. 545, 119627 (2020).
Li, C. et al. A stratified redox model for the Ediacaran ocean. Science 328, 80–83 (2010).
Sahoo, S. K. et al. Ocean oxygenation in the wake of the Marinoan glaciation. Nature 489, 546–549 (2012).
Sperling, E. A., Halverson, G. P., Knoll, A. H., Macdonald, F. A. & Johnston, D. T. A basin redox transect at the dawn of animal life. Earth Planet. Sci. Lett. 371, 143–155 (2013).
Scholz, F. et al. On the isotope composition of reactive iron in marine sediments: redox shuttle versus early diagenesis. Chem. Geol. 389, 48–59 (2014).
Tribovillard, N., Algeo, T., Baudin, F. & Riboulleau, A. Analysis of marine environmental conditions based on molybdenum–uranium covariation—applications to Mesozoic paleoceanography. Chem. Geol. 324, 46–58 (2012).
Calvert, S. E. & Pedersen, T. F. Geochemistry of recent oxic and anoxic marine sediments: implications for the geological record. Mar. Geol. 113, 67–88 (1993).
Algeo, T. J. & Maynard, J. B. Trace-element behavior and redox facies in core shales of Upper Pennsylvanian Kansas-type cyclothems. Chem. Geol. 206, 289–318 (2004).
Scott, C. & Lyons, T. W. Contrasting molybdenum cycling and isotopic properties in euxinic versus non-euxinic sediments and sedimentary rocks: refining the paleoproxies. Chem. Geol. 324-325, 19–27 (2012).
Wang, H. et al. Spatiotemporal redox heterogeneity and transient marine shelf oxygenation in the Mesoproterozoic ocean. Geochim. Cosmochim. Acta 270, 201–217 (2020).
Zhang, K. et al. Oxygenation of the Mesoproterozoic ocean and the evolution of complex eukaryotes. Nat. Geosci. 11, 345–350 (2018).
Gilly, W. F., Beman, J. M., Litvin, S. Y. & Robison, B. H. Oceanographic and biological effects of shoaling of the oxygen minimum zone. Annu. Rev. Mar. Sci. 5, 393–420 (2013).
Schoepfer, S. D. et al. Total organic carbon, organic phosphorus, and biogenic barium fluxes as proxies for paleomarine productivity. Earth Sci. Rev. 149, 23–52 (2015).
Paytan, A. & Griffith, E. M. Marine barite: recorder of variations in ocean export productivity. Deep Sea Res. Part II 54, 687–705 (2007).
Sternberg, E., Tang, D., Ho, T., Jeandel, C. & Morel, F. M. Barium uptake and adsorption in diatoms. Geochim. Cosmochim. Acta 69, 2745–2752 (2005).
Reitz, A., Pfeifer, K., De Lange, G. & Klump, J. Biogenic barium and the detrital Ba/Al ratio: a comparison of their direct and indirect determination. Mar. Geol. 204, 289–300 (2004).
Tyrrell, T. The relative influences of nitrogen and phosphorus on oceanic primary production. Nature 400, 525–531 (1999).
Ozaki, K., Reinhard, C. T. & Tajika, E. A sluggish mid‐Proterozoic biosphere and its effect on Earth’s redox balance. Geobiology 17, 3–11 (2019).
Stacey, J., Hood, A. V. S. & Wallace, M. W. Persistent late Tonian shallow marine anoxia and euxinia. Precambrian Res. 397, 107207 (2023).
Shuster, A. M., Wallace, M. W., van Smeerdijk Hood, A. & Jiang, G. The Tonian Beck Spring Dolomite: marine dolomitization in a shallow, anoxic sea. Sediment. Geol. 368, 83–104 (2018).
Slomp, C. P., Thomson, J. & de Lange, G. J. Controls on phosphorus regeneration and burial during formation of eastern Mediterranean sapropels. Mar. Geol. 203, 141–159 (2004).
Sohm, J. A., Webb, E. A. & Capone, D. G. Emerging patterns of marine nitrogen fixation. Nat. Rev. Microbiol. 9, 499–508 (2011).
Füssel, J. et al. Nitrite oxidation in the Namibian oxygen minimum zone. ISME J. 6, 1200–1209 (2012).
Wang, Z. et al. Coupled nitrate and phosphate availability facilitated the expansion of eukaryotic life at circa 1.56 Ga. J. Geophys. Res. Biogeosci. 125, e2019JG005487 (2020).
Yang, T., Wang, X., Xu, D., Shi, X. & Peng, Y. Nitrogen isotopes from the neoproterozoic liulaobei formation, North China: implications for nitrogen cycling and eukaryotic evolution. J. Earth Sci. 33, 1309–1319 (2022).
Stüeken, E. E. A test of the nitrogen-limitation hypothesis for retarded eukaryote radiation: nitrogen isotopes across a Mesoproterozoic basinal profile. Geochim. Cosmochim. Acta 120, 121–139 (2013).
Acknowledgements
This research was supported by the National Natural Science Foundation of China (42192501, 41921002, and 42130207 to C.Z., 42072036 to W.W., and 42202008 to G.L.), the National Key Research and Development Program of China (2022YFF0800100 to C.Z.), and State Key Laboratory of Palaeobiology and Stratigraphy (20201102 and 213108 to K.P.). We thank Xunlai Yuan for helpful discussions, and Jing Liu and Yangdanjie Zeng for assistance in carbon isotope analysis.
Author information
Authors and Affiliations
Contributions
The study was conceived by C.Z.; Y.S., Y.H., H.S., and X.Z. collected samples; Y.S. and C.G. performed geochemical analyses; Y.S. and C.Z. wrote the paper, with contributions from W.W., X.L., Q.O., K.P., and G.L.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks Samuel Duncanson, and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Sun, Y., Wang, W., Lang, X. et al. A shallow-water oxygen minimum zone in an oligotrophic Tonian basin. Nat Commun 16, 725 (2025). https://doi.org/10.1038/s41467-025-55881-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41467-025-55881-3
