
Abstract
Peripheral neuropathy is a common complication of type 2 diabetes, which is strongly associated with obesity1, causing sensory loss and, in some patients, neuropathic pain2,3. Although the onset and progression of diabetic peripheral neuropathy is linked with dyslipidaemia and hyperglycaemia4, the contribution of inflammation to peripheral neuropathy pathogenesis has not been investigated. Here we used a high-fat, high-fructose diet (HFHFD), which induces obesity and prediabetic metabolic changes, to study the onset of peripheral neuropathy. Mice fed the HFHFD developed persistent heat hypoalgesia after 3 months, but a reduction in epidermal skin nerve fibre density manifested only at 6 months. Using single-cell sequencing, we found that CCR2+ macrophages infiltrate the sciatic nerves of HFHFD-fed mice well before axonal degeneration is detectable. These infiltrating macrophages share gene expression similarities with nerve-crush-induced macrophages5 and express neurodegeneration-associated microglial marker genes6, although there is no axon loss or demyelination. Inhibiting the macrophage recruitment by genetically or pharmacologically blocking CCR2 signalling resulted in more severe heat hypoalgesia and accelerated skin denervation, as did deletion of Lgals3, a gene expressed in recruited macrophages. Recruitment of macrophages into the peripheral nerves of obese prediabetic mice is, therefore, neuroprotective, delaying terminal sensory axon degeneration by means of galectin 3. Potentiating and sustaining early neuroprotective immune responses in patients could slow or prevent peripheral neuropathy.
Similar content being viewed by others

Main
Type 2 diabetes affects millions of individuals and is closely linked with childhood and adult obesity1,7. Approximately 50% of these individuals develop diabetic peripheral neuropathy (DPN)8,9, and about 30–40% of patients with DPN have neuropathic pain, which manifests as spontaneous pain in denervated regions2,4,9,10.
Axonal degeneration in the peripheral nervous system after a traumatic nerve injury has been extensively studied. Following a nerve transection or crush injury, distal axons undergo Wallerian degeneration11,12, and there is a recruitment of immune cells both at the injury site and distally5,13. However, it is unclear whether immune cells contribute to axonal degeneration or disease progression in DPN. To study neuroimmune contributions to early phases of peripheral neuropathy, we modelled diet-induced obesity, prediabetes and metabolic syndrome in mice. The incidence of metabolic syndrome and type 2 diabetes is increasing, owing to the heightened prevalence of obesity and high-caloric food consumption (Western diet)14,15, and many individuals are diagnosed with peripheral neuropathy well before becoming diabetic16. To mimic the consumption of high-caloric food and both boost de novo lipogenesis17,18 and produce obesity, we fed wild-type (WT) male mice on a high-fat diet (60% of kilocalories from lard) supplemented with high-fructose water (42g l−1 d-(−)-fructose; HFHFD; Fig. 1a). This diet leads to both weight gain (Extended Data Fig. 1a) and glucose intolerance (Extended Data Fig. 1b,c,f,g), when compared to a low-fat control diet (CD) supplemented with plain water. In addition, mice fed a HFHFD have a significantly higher level of fasting blood insulin at 12 and 24 weeks than that of CD-fed mice (Extended Data Fig. 1d,h), indicative of insulin resistance, and a significantly higher percentage of glycated HbA1c in blood than that of CD-fed mice (Extended Data Fig. 1e,i), reflecting consistent hyperglycaemia and prediabetes19,20.

a, Illustration of the HFHFD administered to induce prediabetes versus the CD. b, Mechanical withdrawal thresholds of male mice fed a HFHFD or CD over time using an up–down von Frey method (n = 8 mice per group; data are presented as mean values ± s.e.m., P values from two-way analysis of variance (ANOVA) with Sidak multiple comparisons). c, Heat withdrawal latency in Hargreaves assays of male mice fed HFHFD or CD (n = 8 mice per group; data are presented as mean values ± s.e.m., P values from two-way ANOVA with Sidak multiple comparisons). d, Representative heat maps showing the location of a mouse during a 1-h thermal gradient ring recording. e, Time spent on a thermal gradient ring by male mice fed a HFHFD or CD for 12 weeks (n = 8 mice per group; data are presented as mean values ± s.e.m. (shaded area); P = 0.050194 (10 °C), 0.052991 (14 °C), 0.011189 (17 °C), 0.026107 (21 °C), 0.006838 (24 °C), 0.456255 (28 °C), 0.004662 (31 °C), 0.234499 (35 °C), 0.053147 (39 °C), 0.340637 (45 °C), 0.038695 (51 °C) and 0.322922 (56 °C), from multiple unpaired Mann–Whitney tests, *P < 0.05, **P < 0.005). f, Representative PGP9.5 staining of hind paw skin of male mice fed a HFHFD or CD for 24 weeks showing axon terminals in the epidermis marked by PGP9.5 staining and cell layers by 4′,6-diamidino-2-phenylindole (DAPI) staining. Scale bars, 50 μm. g,h, Quantification of IENFD (left) and CGRP+ fibres (right) at 12 weeks of diet (n = 7 (CD) and 8 (HFHFD) for PGP9.5, and n = 9 per group for CGRP; data are presented as mean values ± s.e.m., P values from two-tailed unpaired Student’s t-test; g) and 24 weeks of diet (n = 7 mice per group; data are presented as mean values ± s.e.m., P values from two-tailed unpaired Student’s t-test; h). The graphics in a were created with BioRender.com.
HFHFD leads to peripheral neuropathy
Mice fed the HFHFD develop pain-like behaviour in response to innocuous mechanical stimuli (allodynia) starting at 8 weeks (Fig. 1b), and a reduction in responsiveness to noxious heat (heat hypoalgesia), first detectable at 12 weeks (Fig. 1c). The latter is a prominent feature in patients with diabetic neuropathy21 and in mouse models of diabetes22,23. We evaluated whether there are sex-specific differences in the response to the HFHFD and found that male and female mice gained significantly more weight over time on a HFHFD than on a CD (Extended Data Fig. 2a), and also developed a significant level of mechanical allodynia and heat hypoalgesia (Extended Data Fig. 2b,c). To assess changes in thermal sensitivity, we used an unsupervised thermal gradient ring assay to evaluate thermal preferences and found that mice fed a HFHFD develop a higher tolerance to hot regions of the ring and more cool avoidance (Fig. 1d,e). The mechanical allodynia, unlike the heat hypoalgesia, fully resolved after 24 weeks on the diet, suggesting that the neural dysfunction driving mechanically evoked pain disappears as the disease progresses (Fig. 1b).
Reduction in skin intraepidermal nerve fibre density (IENFD) is an important feature in patients with DPN9. Mice fed a HFHFD do not develop reduced skin IENFD at 12 weeks (Fig. 1g), but terminal axon degeneration is clear after 24 weeks on the diet (Fig. 1h). More specifically, calcitonin gene-related peptide (CGRP)+ fibres in the skin epidermis degenerate in mice at 24 but not 12 weeks, indicating susceptibility of small fibre nociceptor neurons (Fig. 1g,h).
To address whether delayed terminal axon degeneration in HFHFD mice is SARM1 dependent, like Wallerian degeneration11,12,24,25,26, Sarm1-knockout (KO) mice were fed the HFHFD. These mice exhibited the same weight gain as WT mice (Extended Data Fig. 3a,b) but showed a less severe intraepidermal fibre loss in the skin at 24 weeks (Extended Data Fig. 3e). In addition, they did not develop the heat hypoalgesia seen in WT HFHFD mice at 12 weeks (Extended Data Fig. 3c). This suggests that although there is no detectable degeneration at this early stage, the loss of noxious heat sensitivity reflects a SARM1-dependent axonal dysfunction. Sarm1-KO mice did, however, develop mechanical allodynia at 8 weeks on the HFHFD, like WT mice, but this did not resolve at 24 weeks as it does in WT mice (Extended Data Fig. 3d), suggesting that recovery from mechanical allodynia in WT mice at 24 weeks may be a consequence of late mechanosensitive axon terminal degeneration (Extended Data Fig. 3f). Thus, a HFHFD leads to obesity, prediabetic changes and sensory neuropathy, capturing multiple features of clinical type 2 diabetes. We also find that robust behavioural changes are detectable well before axon degeneration, and that the heat sensitivity loss and axonal degeneration in the skin are SARM1 dependent, whereas mechanical allodynia is not.
HFHFD increases nerve macrophages
Diabetes and obesity are accompanied by a metabolic syndrome, and the immune system is implicated in its pathogenesis27. To evaluate whether neuroimmune interactions contribute to peripheral neuropathy, we sought to identify changes in immune cells that may directly affect sensory neurons after 12 weeks on a HFHFD, when mice develop heat hypoalgesia but before axon terminal degeneration is observed. Immune cells reside in close proximity to sensory neurons both in nerves and the skin5,28. To determine whether there are changes in immune cells adjacent to sensory neurons after 12 weeks on the diet, using fluorescence-activated cell sorting, we sorted CD45+ immune cells from the hind paw skin and sciatic nerve, and then we used single-cell sequencing to evaluate immune cell composition (Extended Data Figs. 4a and 8a), analysing 8,462 cells in the sciatic nerve (Extended Data Fig. 4b) and 9,793 cells in the skin (Extended Data Fig. 8b). We identified 13 immune cell clusters in the sciatic nerve. These included five distinct sets of macrophages defined by expression of Csf1r, including two sciatic-nerve-resident macrophage (resMac) populations that did not express Ccr2, of which one expressed Retnla and Clec10a and the other did not express Retnla or Clec10a, and are probably epineural and endoneurial populations5. We also identified a small cluster of resMacs expressing high levels of Cx3cr1, like skin macrophages that contribute to axon growth28. In addition, we identified two dendritic cell clusters that express high levels of MHC class II, H2-Ab1 and Cd74, one of which expresses Cd209a and the other expresses Clec9a and Xcr1, like conventional type 1 dendritic cells. The sciatic nerve also contained T cells of the γδ subtype that express Trdc, innate lymphoid cells and natural killer cells (Extended Data Fig. 4c).
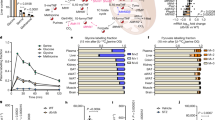
Particularly prominent was the increase in the size of the recruited macrophage (recMac) population in sciatic nerves of mice fed the HFHFD (Fig. 2a,b and Extended Data Fig. 4d). These cells had probably infiltrated recently from the bloodstream as they still expressed monocyte markers (Ccr2, Fn1 and Ly6c2) but had a macrophage identity, expressing Csf1r and Mrc1. We also observed a decrease in the size of the resMac population relative to the increase in the size of the recMacs population. We evaluated the localization of macrophages throughout the nerve in HFHFD-fed mice and found an increase in the number of F4/80+ macrophages in the endoneurial space (Fig. 2c,d). We used flow cytometry to independently validate the increases in size of the recMacs (LY6ChiCD64+CD11B+) population (Fig. 2e and Extended Data Fig. 5). An increase in the number of recMacs was first observed at 12 weeks in both male (Fig. 2e) and female (Extended Data Fig. 2d) mice, which coincides with the onset of heat hypoalgesia, suggesting that macrophages may be recruited in response to changes in neuronal activity or the nerve microenvironment. In addition, we evaluated the relationship of macrophages with the vasculature. We co-stained sciatic nerve sections for a vascular marker, CD31, and saw only a limited association between F4/80+ macrophages and CD31 (Extended Data Fig. 6a). Following perfusion, flow cytometry still revealed an increase in the number of recMacs in the sciatic nerves of mice fed a HFHFD for 12 weeks (Extended Data Fig. 6b). This reveals that macrophages infiltrate the nerve when there is no terminal axonal loss in the skin (Fig. 1g) or demyelination in the nerve (Extended Data Fig. 7a–f), indicating that, quite unlike Wallerian degeneration, macrophage recruitment into diabetic nerves is not driven by axonal degeneration.

a, Uniform manifold approximation and projection (UMAP) plot showing clusters identified in sciatic nerves from mice fed a CD or HFHFD for 12 weeks (2 samples per group). DCs, dendritic cells; ILCs, innate lymphoid cells; snMacs, sciatic nerve macrophages. b, Bar plot of cell proportions showing increase in CCR2+ macrophages in sciatic nerves of HFHFD- versus CD-fed mice. c, Representative cross-section image of sciatic nerves from mice fed a CD or HFHFD for 12 weeks. Col IV, collagen type IV. Scale bars, 100 μm. d, Quantification of F4/80+ macrophages in the endoneurium from nerves of mice fed a CD or HFHFD for 12 weeks (n = 5 mice per group; data are presented as mean values ± s.e.m., P value from two-tailed unpaired Student’s t-test). e, Flow cytometry quantification from sciatic nerves of mice fed a CD or HFHFD for different durations, assessing recMacs (LY6ChiCD64+) in the CD11B+CD45+ live-cell gate (n = 6 mice per group at 8 weeks, 7 mice per group at 10 weeks, and 7 mice in CD and 8 mice in HFHFD at 12 weeks; data are presented as mean values ± s.e.m., P values from two-way ANOVA with Sidak multiple comparisons). f, Volcano plot of gene markers differentially expressed in sciatic nerve resMac (CSF1R+CCR2−) clusters in HFHFD-fed compared to CD-fed mice, highlighting chemokines and cytokines expressed at higher levels; red indicates genes with labels (encoded protein designations) included in the plot. g, Gene Ontology enrichment analysis of significantly (P > 0.05) upregulated genes from resMac clusters; bold indicates chemokine signaling. h, Representative images from sciatic nerves of CCL2–RFP mice fed a CD or HFHFD for 12 weeks. Scale bars, 100 μm. i, RT–qPCR quantification of Ccl2 expression in nerves from mice fed a HFHFD or CD for 8 or 12 weeks (n = 3 mice per group at 8 weeks, and 3 mice in CD group and 4 mice in HFHFD group at 12 weeks; data are presented as mean values ± s.e.m., P values from two-way ANOVA with Sidak multiple comparisons). Arrowheads indicate F4/80+ macrophages that co-express CCL2–RFP; arrows indicate F4/80+ macrophages that do not express CCL2–RFP.
To examine whether there are transcriptional changes in the sciatic nerve resMacs of mice fed a HFHFD, we conducted a differential gene expression analysis between resMac populations in HFHFD versus CD nerves (CSF1R+CCR2− clusters) and found that resMacs in nerves from HFHFD mice upregulate many chemokines, including CCL3 and CCL8 (Fig. 2f). A Gene Ontology pathway analysis revealed that chemotaxis pathways are upregulated in the resMacs (Fig. 2g), suggesting that they recruit recMacs. We fed CCL2–RFP reporter mice with either a CD or HFHFD to determine the source of CCL2 in the nerve. CCL2–RFP was co-localized with the macrophage marker F4/80 in nerves of mice on a HFHFD for 12 weeks (Fig. 2h) but not with neuron or Schwann cell markers (Extended Data Fig. 6c,d), indicating that CCL2 is produced by resMacs. We evaluated the time course of the increase in CCL2 expression in the nerves by quantitative PCR with reverse transcription (RT–qPCR), and this increase was detected at 12 weeks, suggesting that the changes in resMacs that lead to macrophage recruitment occur at this relatively early disease stage (Fig. 2i).
We identified all immune cell types expected in the skin29, including macrophages, dendritic cells, Langerhans cells, T cells, mast cells and innate lymphoid cells (Extended Data Fig. 8c), but found no evidence of immune cell infiltration in the HFHFD mice at 12 weeks (Extended Data Fig. 8c–e). We used flow cytometry to validate these findings with the same gating strategy used for the sciatic nerve (Extended Data Fig. 5) and found no increase in skin recMacs (Extended Data Fig. 8f). Although there was no change in the cellular composition of the skin with the HFHFD, we detected a limited number of differentially expressed genes in the resMac population, indicative not of an inflammatory phenotype but of changes in intracellular signalling (Extended Data Fig. 8g,h). In addition, we evaluated whether there was any change in macrophage proportions near sensory neuron cell bodies in the dorsal root ganglia of mice fed a HFHFD for 12 weeks and found no difference in recMacs proportions compared to those of CD mice (Extended Data Fig. 8i,j). Thus, immune infiltration does not occur in the skin or dorsal root ganglia of mice with diet-induced prediabetes, and macrophage infiltration into nerves is a tissue-specific rather than a systemic immune alteration, probably due to local changes in nerves.
Common nerve immune response
To evaluate whether macrophages recruited into peripheral nerves of mice fed a HFHFD are like those that respond to axonal injury, we compared CSF1R+ clusters obtained from nerves of mice on the HFHFD with a previously reported dataset from sorted sciatic nerve macrophages after a nerve crush injury5 (Fig. 3a). A clustering analysis of the merged datasets revealed that both contain similar cell types, including resMac and recMac clusters. In addition, macrophages from nerves of mice on a CD for 12 weeks and intact nerves showed a similar cellular composition, mainly resMacs (Fig. 3b,d). For recMacs, two clusters emerged in the integrated data, a cluster high in Ccr2 (CCR2+ recMacs) and a cluster high in Lgals3 (LGALS3hi recMacs; Fig. 3d) expression in both HFHFD (12 weeks) and post-crush (day 1 and day 5) nerve samples (Fig. 3d). The recMacs in both the HFHFD and injured nerve samples also express neurodegenerative-disease-associated microglial6 gene markers such as Trem2 and Lgals3 (Fig. 3f,g).

a, Integration of macrophages from our HFHFD dataset and a published nerve crush dataset5. b, Cell proportions under the different conditions in the integrated datasets. c, Principal component (PC) analysis plot of cell proportions. d, UMAP plots of the merged dataset split by condition. e, Volcano plot of gene markers differentially expressed between CCR2+ macrophages (recMacs) and CCR2− macrophages (resMacs). f,g, Violin plots of expression levels of Lgals3 (f) and Trem2 (g) in the merged macrophage datasets. h, Representative flow cytometry plots (left) of recMacs (RMs) (CD64+LY6C+) from the crushed (ipsilateral) and control (contralateral) sciatic nerves pre-gated on live CD45+CD11B+LY6G− myeloid cells with quantification (right; n = 4 mice per group; data are presented as mean values ± s.e.m., P value from two-tailed unpaired Student’s t-test). Numbers in outlined areas (left) indicate percent cells in each. i,j, RT–qPCR quantification of Lgals3 (i) and Trem2 (j) expression from nerves of mice fed a CD or HFHFD for 12 weeks or CD mice 1 day post sciatic nerve crush (n = 3 mice (CD), 5 mice (HFHFD) and 4 mice (crush group); data are presented as mean values ± s.e.m., P values from Brown–Forsythe and Welch one-way ANOVA, Dunnett T3 multiple comparison test). The graphics in a were created with BioRender.com.
We identified many genes differentially expressed between recMac and resMac populations in the whole dataset, including genes encoding injury-responsive molecules, such as Fn1 and Chil35, as well as genes encoding molecules responsible for lipid uptake, such as Lpl30,31,32,33, and genes encoding molecules that promote nerve regeneration, including Lgals3 and Thbs133,34,35 (Fig. 3e). To validate these findings, we performed a sciatic nerve crush and found that the size of the population of recMacs detected by flow cytometry in HFHFD-fed mice also increased in crushed nerves (Fig. 3h), when the same gating strategy was used (Extended Data Fig. 5). In addition, using RT–qPCR, we found that both nerve crush and a HFHFD lead to increased expression of Lgals3 and Trem2, albeit at different magnitudes (Fig. 3i,j). Macrophages that infiltrate intact nerves of mice fed a HFHFD are similar, therefore, to those recruited rapidly and distally after a traumatic nerve injury, even though the two insults are quite distinct, and both express pro-repair genes.
Macrophages delay onset of neuropathy
recMacs in injured peripheral nerves contribute to the clearance of both myelin and axonal debris, enabling axonal regeneration and recovery of function36. To evaluate whether macrophages recruited into nerves of mice fed a HFHFD have a neuroprotective role, we fed Ccr2-KO and WT mice on the HFHFD and evaluated whether preventing CCR2-dependent macrophage recruitment into nerves had any impact on axonal disease progression (Fig. 4a). The number of LY6ChiCd64+ recMacs in the sciatic nerve of Ccr2-KO mice on a HFHFD was reduced compared to that of WT mice, indicating that macrophage recruitment in HFHFD mice is CCR2 dependent (Fig. 4b). The number of T cells did not decrease (Extended Data Fig. 9f). Ccr2-KO mice on the HFHFD exhibited a greater heat hypoalgesia than age-matched WT mice (Fig. 4c), but no difference in mechanical allodynia (Fig. 4g). Ccr2-KO mice at 12 weeks on the HFHFD also had a reduced IENFD, revealing a much earlier onset of terminal axonal neuropathy than in WT mice on the HFHFD (3 rather than 6 months; Fig. 4d,e), but by 6 months, both WT mice and Ccr2-KO mice exhibited similar IENFD loss and heat hypoalgesia (Fig. 4g and Extended Data Fig. 9h). The Ccr2-KO mice fed the HFHFD for 12 weeks gained weight to the same extent as their WT counterparts (Extended Data Fig. 9a) and exhibited the same glucose intolerance as WT mice at 12 weeks of HFHFD feeding (Extended Data Fig. 9b,c). Although Ccr2-KO mice had lower fasting insulin levels than WT mice, as reported previously37 (Extended Data Fig. 9e), they had glycated HbA1c levels similar to those of WT mice after HFHFD feeding for 12 weeks (Extended Data Fig. 9d), suggesting a similar prediabetic status with consistent hyperglycaemia.

a, Illustration of experiment. b, Quantification of flow cytometry data of recMacs (LY6C+Cd64+CD11B+) in sciatic nerves of male Ccr2-KO and WT mice fed the HFHFD for 12 weeks (n = 5 mice per group; data are presented as mean values ± s.e.m., P value from two-tailed unpaired Student’s t-test). c, Heat withdrawal latencies in Hargreaves assays of male Ccr2-KO and WT mice fed HFHFD (n = 5 mice per group; data are presented as mean values ± s.e.m., P values from two-way ANOVA with Sidak multiple comparisons). d, Representative PGP9.5 staining of hind paw skin (n = 5 mice per group; scale bars, 50 μm). e, Quantification of IENFD of male Ccr2-KO and WT mice fed a HFHFD for 12 or 24 weeks (n = 5 mice per group; data are presented as mean values ± s.e.m., P values from two-way ANOVA with Sidak multiple comparisons). f, Timeline of CCR2 and CCR5 inhibitor, CVC, injections starting 8 weeks on HFHFD. g, Heat withdrawal latencies in Hargreaves assays of CVC- and vehicle-injected male mice fed HFHFD (n = 9 mice (vehicle) and 8 mice (CVC) group; data are presented as mean values ± s.e.m., P values from two-way ANOVA with Sidak multiple comparisons). h, Quantification of IENFD of CVC- and vehicle-injected male mice fed HFHFD at 12 weeks (n = 6 mice (vehicle) and 4 mice (CVC); data are presented as mean values ± s.e.m., P value from two-tailed unpaired Student’s t-test). i, Illustration of experiment. j, Heat withdrawal latencies in Hargreaves assays of WT and Lgals3-KO mice fed HFHFD for 12 weeks (n = 10 mice per group; data are presented as mean values ± s.e.m., P value from two-tailed unpaired Student’s t-test). k, Quantification of IENFD of WT and Lgals3-KO mice fed HFHFD for 12 weeks (n = 9 mice (CD) and 10 mice (HFHFD); data are presented as mean values ± s.e.m., P value from two-tailed unpaired Student’s t-test). The graphics in a,i were created with BioRender.com.
To block CCR2 recruitment after onset of the disease, we used the CCR2 and CCR5 inhibitor cenicriviroc (CVC)38. After 8 weeks on a HFHFD, mice were randomly placed on CVC treatment or vehicle (Fig. 4f). Mice treated with CVC that did not lose weight were evaluated for behavioural and histological changes (Extended Data Fig. 10a). Like the Ccr2-KO mice, the CVC-injected mice had persistent mechanical allodynia (Extended Data Fig. 10b) and higher heat latencies than the vehicle-treated group at 12 weeks on the HFHFD (Fig. 4g). They also had detectable axonal loss in the epidermis at 12 weeks, consistent with earlier peripheral axonal disease onset, as with Ccr2-KO mice (Fig. 4h). Mice fed with the CD and given CVC or vehicle injections between 8 and 12 weeks did not exhibit changes in heat sensitivity or IENFD (Extended Data Fig. 10c,d). We also evaluated temperature preferences in both female and male HFHFD-fed mice following the macrophage recruitment blockade strategies and found that both male and female mice lacking CCR2 signalling had more heat tolerance on the thermal gradient ring than control counterparts, confirming diminished heat sensitivity (Extended Data Fig. 10e,f).
One of the most upregulated genes in CCR2+ macrophages recruited into nerves after HFHFD exposure was Lgals3 (Fig. 3e). Lgals3 encodes galectin-3 (Gal-3), a β-galactoside-binding lectin, which can be intracellular, membrane-bound or secreted by macrophages39. Gal-3 is expressed by microglia at sites of brain atrophy and is protective against retinal degeneration35. We evaluated whether knockdown of Lgals3 leads to earlier neuropathy onset (Fig. 4i). After 12 weeks of HFHFD, Lgals3-KO mice gained the same amount of weight as WT mice (Extended Data Fig. 10g) but had more severe heat hypoalgesia (Fig. 4j), as well as detectable IENFD loss, unlike WT mice (Fig. 4k). We also used flow cytometry to evaluate the effect of Lgals3 knockdown on macrophage recruitment and found that Lgals3-KO nerves had recMac recruitment similar to that of WT nerves after 12 weeks of the HFHFD (Extended Data Fig. 10h). CCR2-dependent recruitment of macrophages into peripheral nerves of mice fed a HFHFD therefore both reduces the severity of behavioural sensory loss and delays the progression of axonal degeneration in the skin by means of Gal-3 (Extended Data Fig. 10i). Blocking recruitment of macrophages into peripheral nerves in obese prediabetic mice is therefore detrimental, whereas promoting and sustaining this recruitment or signalling by means of Gal-3 is beneficial.
Discussion
In a diet-induced model of obesity and prediabetes, macrophages with an expression profile resembling that of neurodegenerative-disease-associated myeloid cells infiltrate into intact peripheral nerves, and this recruitment is neuroprotective, delaying the onset of terminal axonal degeneration by means of Gal-3. In the HFHFD model, mechanical allodynia developed early in the disease (2 months) but resolved by 6 months. Transient mechanical allodynia is also observed in a genetic model of leptin-deficiency-induced obesity (db/db)40. Preventing axonal degeneration by Sarm1 KO caused persistence of the mechanical allodynia, suggesting that blocking axon degeneration does not eliminate pain hypersensitivity. Heat hypoalgesia manifested in HFHFD-treated mice well before axon terminal degeneration was detectable, but Sarm1 KO prevented the lack of sensitivity to noxious heat even before axon degeneration occurs. Individuals with prediabetes often experience sensory abnormalities years before a definitive skin biopsy diagnosis of axonal loss16, suggesting that the HFHFD prediabetic mouse model captures major features of the human disease21,41. The mechanism responsible for the early SARM1-dependent pre-neuropathy heat hypoalgesia remains to be determined.
Macrophages are recruited into peripheral nerves but not into the skin of prediabetic mice. Mice on a high-fat diet also develop an upregulation of Lgals3 in the sciatic nerve42, a gene expressed at high levels in those recMacs that infiltrate into the nerves of HFHFD diabetic mice (Fig. 4f). Knockdown of Gal-3 delays axonal regeneration after nerve injury32,34 and is a TREM2 ligand in microglia reacting to neurodegeneration43. Gal-3-expressing phagocytic macrophages infiltrate into diabetic brains44, suggesting that peripheral-nerve-infiltrating macrophages may be phagocytic and neuroprotective by virtue of clearing debris, excess fatty acids or metabolites such as deoxysphingolipids, which accumulate in nerves22,33. Mice lacking Gal-3 exhibit more severe heat hypoalgesia and earlier sensory neuron retraction in the skin, indicating that Gal-3 is protective against sensory axon degeneration in peripheral neuropathy. Gal-3 recognizes advanced glycation end (AGE) products and promotes their phagocytosis and clearance45, and AGEs accumulate in DPN46,47. Gal-3 acts in an opposite manner from RAGE, the canonical AGE receptor, which is detrimental in DPN48, suggesting that both protective and detrimental immune responses coexist in nerves of mice with peripheral neuropathy. Thrombospondin 1, another neuroprotective mediator33, is expressed at high levels by nerve-infiltrating macrophages and inhibits the peripheral sensitization of nociceptors29. Macrophages infiltrating the nerves of HFHFD-fed mice express high levels of Trem2, which encodes a molecule involved in microglial neuroprotective responses to neurodegeneration, and TREM2 variants that decrease TREM2 signalling increase degeneration49,50,51. Whether TREM2 gene variants impact human diabetic neuropathy needs to be investigated. The similarities between recMacs in prediabetic peripheral nerves and after a nerve injury, as well as in disease-associated microglia6, reveal a common myeloid cell identity that may reflect similar neuroprotective roles in different disease states.
Macrophages may be recruited into peripheral nerves during prediabetes in response to changes in the nerve microenvironment. Non-neuronal cell types, such as Schwann cells, may be modified by the HFHFD, as they are responsive to changes in glucose or insulin42 and may release mediators that alter nerve-resident macrophages to trigger recruitment of circulating myeloid cells. Metabolomic studies show that a high-fat diet alone and when supplemented with low-dose streptozotocin modifies the fatty acid composition of the nerve52. It is possible that 12 weeks on a HFHFD results in an excess of free fatty acids in nerves that triggers resMacs in a manner similar to membrane or myelin debris in acutely traumatized nerves. Macrophages could also be recruited by Gal-3 to clear AGE accumulation in the nerve46,47. Excess AGEs may prevent CCR2+ macrophages from maintaining constant clearance, promoting degeneration at late time points. It is also plausible that macrophages exert neuroprotective effects by signalling to Schwann cells, as in traumatic nerve injury53.
Although macrophages have long been considered to have a pathogenic role in diabetes and insulin resistance54,55, we find that macrophages recruited to nerves early in the disease delay the onset of peripheral neuropathy and reduce its extent. An increase in repair-like macrophages in RAGE-null mice also alleviates type 1 diabetic DPN48. However, that study was carried out in a streptozotocin injection model with no changes in IENFD48. Although macrophages were suggested to play a detrimental role in a high-fat-diet-induced model of neuropathy56,57, that study did not differentiate between macrophage subtypes in the nerve and did not investigate heat sensitivity or skin innervation, only mechanical allodynia. Mechanical allodynia was transient in our HFHFD mouse model, and its resolution may be driven by the degeneration of terminal mechanosensitive fibres, as supported by our Sarm1-KO results. In addition, mechanical allodynia is the least consistent symptom in patients with diabetes, unlike sensory loss and axon terminal loss in the skin4,21, and mouse models need to capture these features of the disease. As the transcriptional profile of macrophages recruited to prediabetic nerves is similar to that of macrophages in crushed nerves, and because their role is neuroprotective, it will be important to determine whether this protection also occurs in non-metabolic peripheral neuropathies, such as chemotherapy-induced neuropathy. Our findings reveal a neuroprotective role of immune cells in a diet-induced peripheral neuropathy that can now drive exploration of biomarkers that may predict the risk of neuropathy, as well as therapeutic strategies that maximize and sustain the neuroprotective action of macrophages in nerves of individuals at high risk.
Methods
Mice
C57BL/6J mice 8–12 weeks of age were obtained from the Jackson Laboratory (JAX: 000664). Ccr2-KO (strain number 004999), Sarm1-KO (strain number 018069), CCL2–RFP (strain number 016849) and Lgals3-KO (strain number 006338) mice were obtained from the Jackson Laboratory. All animal experiments and procedures were conducted according to the institutional animal care and safety guidelines at Boston Children’s Hospital. All mice in this study were kept on a 12 h light cycle, at 21–23 °C, with 30–50% humidity.
Diet
A high-fat diet containing 60% fat from lard was obtained from Research Diets (D12492), along with a matched sucrose CD (D12450J). Diet experiments were started at 10–12 weeks of age. Whole cages of mice were randomly assigned to receive the CD or HFHFD. Mice that were given the HFHFD were also given water bottles with 42 g l−1 d-(−)-fructose in autoclaved water, and those given CD were given plain autoclaved water. Mouse food and water levels were monitored three times per week and replenished as needed throughout the entirety of their diet regimen.
Glucose tolerance test
Mice were fasted overnight, and baseline blood glucose was measured from a tail nick using a One Touch Ultra 2 glucose monitoring system. Mice were then injected with 5 μl g−1 body weight of 0.2 g ml−1 glucose i.p., and blood glucose was measured at 15, 30, 60 and 120 min.
HbA1c assay
Mice were fasted for 4 h, and whole blood was collected using cardiac puncture with EDTA and stored at −20 °C. The Mouse Hemoglobin A1c assay kit (Crystal Chem, catalogue number 80310) was used to measure mouse glycated haemoglobin levels following the manufacturer’s protocol. In brief, blood samples were lysed using the provided lysis buffer and the absorbance was measured at 700 nm twice following two enzymatic protease digestions. The provided calibrators were used to calculate the percentage of glycated HbA1c in each sample.
Insulin enzyme-linked immunosorbent assay
Mice were fasted for 4 h, and whole blood was collected using cardiac puncture with EDTA. Blood was then centrifuged at 4 °C for 20 min at 2,000g to collect plasma, which was then frozen at −20 °C until the assay was carried out. Insulin levels in each sample were measured using the Ultrasensitive Mouse Insulin ELISA kit (Crystal Chem, catalogue number 90080) following the manufacturer’s protocol for the wide-range assay. In brief, samples and standard solutions were incubated in the pre-coated enzyme-linked immunosorbent assay wells provided. The plate was then washed and incubated in an anti-insulin-conjugated solution followed by washes and substrate solution incubation. Finally, the reaction was stopped, and the absorbance was measured at 450 nm and 630 nm and calculated using the standard curve.
Behaviour
von Frey
Mice were habituated to von Frey chambers and mesh bottoms for 2 days, 1 h each day, before testing. On the day of the test, mice were habituated for 1 h before the test. Mice were placed in behaviour chambers in random order at every timepoint as a mean of blinding investigators, and there was no consistent placement for each mouse. The up–down method was used to determine the mice’s 50% withdrawal threshold58. Behavioural experiments to compare CD and HFHFD were otherwise difficult to conduct in a blinded fashion, as mouse weights and faeces colours were obvious; however, these were conducted by at least two independent investigators to validate the findings. All subsequent experiments with genetic or pharmacological manipulations were conducted in a blinded fashion (as was the Hargreaves test).
Hargreaves
Mice were habituated to the Hargreaves apparatus (IITC number 390G) consisting of a glass floor heated to 30 °C and a Plexiglass chamber for 2 days before testing, 30 min each day. On the day of the assessment, mice were habituated for 1 h before the test. A focused heat light source was shined on the plantar surface of the left paw of the mice, and a ramping heat stimulus was therefore applied until paw withdrawal was recorded, with a time limit of 30 s. Readings were averaged from three trials per mouse.
Thermal gradient ring
Mice were placed in the thermal gradient ring (Ugo Basile, 35550), which was set to 10 °C on one side and 56 °C on the other side. Their location and movement and the temperature of the plates were recorded for an hour. All mice in a single experiment were recorded using one apparatus. Using the ANY MAZE software, the time spent per zone and distance travelled were recorded for each mouse.
Skin immunohistochemistry and IENFD analysis
A 2-mm punch from the hind paw plantar skin of mice was collected and fixed in Zamboni Fixative (Newcomer, number 1459A) overnight at 4 °C on a piece of filter paper to flatten the tissue. After fixation, skin samples were kept in 30% sucrose for at least 48 h in 4 °C. Skin samples were then frozen in OCT, cryosectioned at 30 μm thickness and mounted onto SuperFrost Plus microscope slides. For immunostaining, slides were thawed for 1 h at room temperature and then washed in PBS for 15 min following three 10-min 1% Triton-X washes. Slides were incubated in blocking buffer for 2 h at room temperature (10% donkey serum, 0.4% Triton-X, 0.05% Tween 20, 1% BSA) and then incubated in primary antibody overnight at 4 °C (rabbit anti-PGP9.5 Abcam catalogue number ab108986, 1:200). The next day, slides were washed three times with PBS and incubated in secondary antibody for 2 h at room temperature (donkey anti-rabbit Cy3 Jackson ImmunoResearch 711-165-152 1:500). Slides were then washed three times with PBS and mounted with Prolong Antifade DAPI medium (Invitrogen, number P36935). Confocal images were taken on a Leica SP8 microscope with a ×63 oil objective. A z-stack spanning the 30 μm thickness was taken and four adjacent images were tiled. Using FIJI (ImageJ), analysis to determine IENFD was conducted by an investigator blinded to the treatment groups. Maximum-intensity projection was taken, and the number of fibres crossing the dermo-epidermal boundary was counted and divided by the length of the epidermal layer in the image to obtain an IENFD value for that image following previously described rules59. At least three images of non-consecutive sections were averaged per mouse to obtain the IENFD value for that mouse.
Sciatic nerve tissue collection and processing
Sciatic nerves were dissected and collected in ice-cold 1% BSA in RPMI. For digestion, tissues were placed in collagenase A (Roche, 5 mg ml−1) and Dispase II (Roche, 1 mg ml−1) and were minced before being moved to a thermomixer at 37 °C for 1 h at 1,000 r.p.m. Samples were then filtered using a 70-μm filter, washed in FACS buffer (2% fetal bovine serum (FBS), 2 mM EDTA, PBS) and then washed with 1× PBS to obtain single-cell suspensions.
Skin tissue collection and processing
Skin punch biopsies were dissected and collected in ice-cold 1% BSA in RPMI. Tissues were minced and Liberase (Sigma-Aldrich, catalogue number 05401119001) was added at a final concentration of 1 mg ml−1, and tubes were placed on a thermomixer at 37 °C for 1.5 h at 1,000 r.p.m. Samples were then filtered using a 70-μm filter, washed in FACS buffer (2% FBS, 2 mM EDTA, PBS) and then washed with 1× PBS to obtain single-cell suspensions.
Dorsal root ganglion collection and processing
Lumbar dorsal root ganglia were collected in ice-cold Dulbecco’s modified Eagle medium (DMEM) containing FBS plus penicillin and streptomycin. Dorsal root ganglia were digested in collagenase A (Roche, 5 mg ml−1) and Dispase II (Roche, 1 mg ml−1) for 70 min. Digested dorsal root ganglia were triturated using large-, medium- and small-sized polished glass pipettes in DMEM containing DNase. The cells were resuspended in DMEM containing FBS plus penicillin and streptomycin and were overlaid on 10% BSA solution. The bilayer was centrifuged for 12 min at 1,000g at reduced acceleration and deceleration. The top two layers were discarded, and the cell pellet was resuspended in FACS buffer (2% FBS, 2 mM EDTA, PBS) and then washed with 1× PBS to obtain single-cell suspensions.
Flow cytometry
Cells were then incubated for 10 min at 4 °C in Fc Block (Tonbo Biosciences, catalogue number 70-0161-U500, 1:2,000) and then washed with FACS buffer. Samples were then stained for 30 min with antibodies to mouse CD45–ef780 (Thermo Fisher Scientific, clone 30-11, catalogue number 47-0451-82, 1:400), CD11B–FITC (Thermo Fisher Scientific, clone M1/70, catalogue number 11-0112-82, 1:400), CD64–PE-594 (BioLegend, clone X54-5/7.1, catalogue number 139319, 1:400), LY6C–BV711 (BioLegend, clone HK1.4, catalogue number 128037, 1:1,000), LY6G–PE (BioLegend, clone 1A8, catalogue number 127607, 1:800), CD3ε–PE (BioLegend, clone 145-2C11, catalogue number 100307, 1:400) for 30 min on ice and then washed with FACS buffer twice and resuspended with 3 μM DAPI before being run on an LSR Fortessa Cytometer using BDFACS Diva Software and analysis using FlowJo.
Preparation of immune cells for single-cell sequencing
Single-cell suspensions collected as described above from two mice (four sciatic nerves) were pooled in each sample. Samples were resuspended in FACS buffer (2% FBS, 2 mM EDTA, PBS) and then incubated for 10 min at 4 °C in Fc Block (Tonbo Biosciences catalogue number 70-0161-U500, 1:2,000) and washed with FACS buffer. Cells were then stained for 30 min in anti-CD45 conjugated APC–ef780 antibody (Thermo Fisher Scientific, clone 30-11, catalogue number 47-0451-82, 1:200) in FACS buffer and then washed twice in PBS with 0.5% BSA and no EDTA. Cells were then resuspended with 3 μM DAPI in PBS with 0.5% BSA before live CD45+ cells were sorted on a BD FACS Aria sorter using BDFACS Diva Software and processed for 10× sequencing following a 10× protocol.
Barcoding and library preparation
The Chromium Next GEM Single Cell 3′ Reagent kit v3.1 (Dual Index) was used. Barcoding and library preparation were performed following the manufacturer’s protocols. In brief, to generate single-cell gel-bead-in-emulsion solution, sorted cells were resuspended in a final volume of 40 μl and were loaded on a Next GEM Chip G (10X Genomics) and processed with the 10X Genomics Chromium Controller. Reverse transcription was performed as follows: 53 °C for 45 min and 85 °C for 5 min in a thermocycler. Next, first-strand cDNA was cleaned with DynaBeads MyOne SILANE (10X Genomics, 2000048). The amplified cDNAs, intermediate products and final libraries were prepared and cleaned with the SPRIselect Regent kit (Beckman Coulter, B23318), and examined on a High-Sensitivity DNA Chip with the Bioanalyzer system (Agilent). Ten-microlitre aliquots of cDNAs were use for library preparation following the manufacturer’s instructions (10X Genomics). The final libraries were examined on High-Sensitivity DNA Tape with TapeStation (Agilent) before pooling for sequencing. A small aliquot of each library was used for quality control to determine fragment size distribution and DNA concentration using a bioanalyser. Libraries were pooled for sequencing with a NovaSeq 6000 (Illumina) at an estimated depth of 113,000 reads per cell in the nerve and 65,000 reads per cell in the skin. Sequencing reads were aligned to the mouse reference transcriptome (mm10, version 2020-A) using CellRanger (v7.0.1).
Single-cell RNA-sequencing data analysis
Raw single-cell RNA-sequencing data were processed using 10X Genomics CellRanger software version 7.0.01. The CellRanger mkfastq function was used for de-multiplexing and generating FASTQ files from raw BCL. The CellRanger count function with default settings was used with the mm10 reference supplied by 10X Genomics, to align reads and generate single-cell feature counts. Ambient RNA contamination was removed from each sample using CellBender6 (remove-background, default parameters). Seurat version 5.0.1 implemented in R version 4.3.2 was used for downstream analysis. Cells were excluded if they had fewer than 500 features, or fewer than 250 genes, or the mitochondrial content was more than 20%. Two CD and two HFHFD samples were integrated and normalized following a previously described Seurat SCTransform+ CCA integration pipeline60. The mitochondrial mapping percentage was regressed out during the SCTransform normalization step. PCA was performed and the top 40 principal components were used for downstream analysis. A K-nearest-neighbour graph was produced using Euclidean distances. The Louvain algorithm was used with the resolution set to 0.4 to group cells together. Nonlinear dimensional reduction was carried out using UMAP. The FindAllMarkers function was used to determine marker genes for each cluster and clusters were identified on the basis of canonical markers previously published in the literature5,13,29. Differentially expressed genes were determined using the FindMarkers function that performs differential expression testing based on the non-parametric Wilcoxon rank sum test.
Integration with macrophages after nerve crush dataset
All clusters expressing Csf1r were subset into a new dataset and raw data file from ref. 5 were downloaded. Samples from two CD mice and two HFHFD mice were merged using Seurat with Ydens’ naive, day 1 post-crush and day 5 post-crush samples. Data were filtered according to the same filters above and integrated using SCTransform as described above. The dataset was then re-clustered with 0.4 resolution, and FindAllMarkers was used to identify cell types.
Sciatic nerve immunohistochemistry
Mice were anaesthetized after 12 weeks of CD or HFHFD feeding using 200 mg kg−1 pentobarbital and perfused transcardially using PBS. Sciatic nerves were collected in 4% paraformaldehyde and left in 4 °C overnight and then moved to 30% sucrose solution. Nerves were then frozen in OCT, cryosectioned as 50-μm cross-sections and mounted onto SuperFrost Plus microscope slides. For immunostaining, slides were thawed for 1 h at room temperature and then washed in PBS for 15 min following three 10-min 1% Triton-X washes. Slides were incubated in blocking buffer for 2 h at room temperature (10% donkey serum, 0.4% Triton-X, 0.05% Tween 20, 1% BSA) and then incubated in primary antibody overnight at 4 °C: rat anti-F4/80 (Bio-Rad, catalogue number MCA497GA, 1:250), goat anti-collagen type IV (Sigma-Aldrich, catalogue number AB769, 1:500), goat anti-CD31 (Novus Biologicals, catalogue number AF3628, 1:25). The next day, slides were washed three times with PBS and incubated in secondary antibody for 2 h at room temperature: donkey anti-rat 488 (Abcam, catalogue number ab150153, 1:500), donkey anti-goat 647 (Invitrogen, catalogue number A21447, 1:500). Slides were then washed three times with PBS and mounted with Prolong Antifade DAPI medium (Invitrogen, catalogue number P36935). Confocal images were taken on a Leica SP8 microscope with a ×20 air objective. A z-stack spanning the whole thickness was taken. A maximum-intensity projection was generated using ImageJ, and the area within the epineurium (collagen staining) was measured. The number of F4/80+ macrophages associated with a nucleus was counted and divided by the area of the endoneurial space in each image.
Nerve electron microscopy
Sciatic nerves were collected in 2.5% glutaraldehyde, 2.5% paraformaldehyde in 0.1 M sodium cacodylate buffer (pH 7.5) and fixed in the same solution overnight. Small pieces (1–2 mm) of fixed tissue were washed in 0.1 M cacodylate buffer and postfixed with 1% osmium tetroxide (OsO4) and 1.5% potassium ferrocyanide (KFeCN6) for 1 h, washed in water twice, then washed once in 50 mM maleate buffer pH 5.15 (MB) and incubated in 1% uranyl acetate in MB for 1 h followed by one wash in MB, two washes in water and subsequent dehydration in grades of alcohol (10 min each; 50%, 70%, 90%, 2 × 10 min 100%). The samples were then placed in propyleneoxide for 1 h and infiltrated overnight in a 1:1 mixture of propyleneoxide and TAAB Epon (TAAB Laboratories Equipment). The next day, samples were embedded in TAAB Epon and polymerized at 60 °C for 48 h. Ultrathin sections were cut on a Reichert Ultracut-S microtome, picked up on copper grids, stained with lead citrate and examined in a JEOL 1200EX Transmission electron microscope or a TecnaiG2 Spirit BioTWIN. Images were recorded with an AMT 2k CCD (charge-coupled device) camera. Electron microscopy imaging, consultation and services were performed at the Electron Microscopy Facility at Harvard Medical School. g ratios and percentages of myelinated axons were calculated from the EM images using MyelTracer as described before61.
Mouse injections
The CCR2 and CCR5 inhibitor CVC (Selleckchem, catalogue number S8512) or dimethylsulfoxide was diluted in corn oil and injected i.p. at 20 mg kg−1 three times per week starting 8 weeks on the HFHFD until 12 weeks. Mice were chosen randomly from each cage. Mouse weights were recorded weekly, and two mice in each group that lost weight as a result of the drug treatment were excluded from the study before proceeding with behavioural testing because this counteracted the weight gain expected from HFHFD feeding. For behavioural testing, the person injecting mice or placing mice in behaviour chambers was different from the person investigating the behaviour, who was therefore fully blinded.
Sciatic nerve crush surgery
Mice were anaesthetized by administering 2.5% isoflurane. A sciatic nerve crush was performed by exposing the left sciatic nerve at the mid-thigh level and crushing it with a haemostat at the second stop for 30 s. The surgical incision was then closed using silk sutures and the mouse was monitored every day after the surgery.
RT–qPCR
Mouse sciatic nerves were collected in RNAlater and stored at 4 °C overnight before long-term storage at −80 °C. To isolate RNA, nerves were moved from RNAlater to RLT buffer (Qiagen) and minced. A handheld homogenizer was then used to homogenize the nerves in RLT buffer, and an equal volume of 70% ethanol was then added to the tube. RNA was isolated following the Qiagen mini kit protocol. cDNA was synthesized from the resulting RNA using the cDNA VILO kit (Invitrogen, catalogue number 11754050). Relative gene expression was determined using gene-specific primers (PrimerBank) and SYBR Green master mix (Life Technologies) on a 7500 Fast Real-time PCR system (Applied Biosystems). Expression levels were normalized to Hprt levels using the 2−ΔCt method. The following primer sequences were used—Hprt forward: 5′-CAGTCCCAGCGTCGTGATTA-3′, Hprt reverse: 5′-TGGCCTCCCATCTCCTTCAT-3′; Ccl2 forward: 5′-GATGCAGTTAACGCCCCACT-3′, Ccl2 reverse: 5′-GAGCTTGGTGACAAAAACTACAGC-3′, Lgals3 forward: 5′-TAATCAGGTGAGCGGCACAG-3′, Lgals3 reverse: 5′-ATAGGGCACCGTCAGTGGTC-3′, Trem2 forward: 5′-ATGGGACCTCTCCACCAGTT-3′, Trem2 reverse: 5′-CACAGGATGAAACCTGCCTGG-3′.
Statistical analysis
Sample size for behavioural experiments was based on previous laboratory data and data in the literature62,63,64. Sample size for the single-cell RNA-sequencing experiment was based on common practice at n = 2 for single-cell transcriptomics. We also utilized an available tool to estimate sample size65 using our knowledge of the frequency of rare populations (for example, neutrophils in healthy tissue), and determined that the number of cells in our analyses was above that needed for 95% probability of success at detecting rare populations. Statistical analysis was performed using GraphPad Prism software. For each experiment of its kind, we conducted a normality test using the Shapiro–Wilk test, and they all passed the normality test; therefore, we assumed normality for subsequent tests from the same datasets. This was the case for body weight, von Frey, Hargreaves, flow cytometry, IENFD, fasting insulin, glucose tolerance test, HbA1c and nerve immunohistochemistry data. For statistical comparison between groups, two-group comparisons were made using two-tailed unpaired Student’s t-test. For comparisons of two groups over time, two-way ANOVA with appropriate multiple comparisons tests was used, and 0.05 was set as the threshold for significance. Plots from behaviour assays, immunohistochemistry and flow cytometry are representative of at least two independent repetitions. All error bars plotted represent s.e.m.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The single-cell sequencing raw files have been deposited in the Gene Expression Omnibus database under the accession number GSE255553. The single-cell sequencing dataset from the sciatic nerve crush is available at the Gene Expression Omnibus under the accession number GSE144708. The mouse reference transcriptome (mm10, version 2020-A) was used for analysis. Source data are provided with this paper.
References
Bacha, F. et al. Pathophysiology and treatment of prediabetes and type 2 diabetes in youth. Diabetes Care https://doi.org/10.2337/DCI24-0029 (2024).
Feldman, E. L. et al. Diabetic neuropathy. Nat. Rev. Dis. Primers 5, 41 (2019).
von Hehn, C. A., Baron, R. & Woolf, C. J. Deconstructing the neuropathic pain phenotype to reveal neural mechanisms. Neuron 73, 638–652 (2012).
Eid, S. A. et al. New perspectives in diabetic neuropathy. Neuron 111, 2623–2641 (2023).
Ydens, E. et al. Profiling peripheral nerve macrophages reveals two macrophage subsets with distinct localization, transcriptome and response to injury. Nat. Neurosci. 23, 676–689 (2020).
Deczkowska, A. et al. Disease-associated microglia: a universal immune sensor of neurodegeneration. Cell 173, 1073–1081 (2018).
Suzuki, K. et al. Genetic drivers of heterogeneity in type 2 diabetes pathophysiology. Nature 627, 347–357 (2024).
Sun, H. et al. IDF Diabetes Atlas: global, regional and country-level diabetes prevalence estimates for 2021 and projections for 2045. Diabetes Res. Clin. Pract. https://doi.org/10.1016/j.diabres.2021.109119 (2021).
Jensen, T. S. et al. Painful and non-painful diabetic neuropathy, diagnostic challenges and implications for future management. Brain 144, 1632–1645 (2021).
Elafros, M. A. et al. Towards prevention of diabetic peripheral neuropathy: clinical presentation, pathogenesis, and new treatments. Lancet Neurol. 21, 922–936 (2022).
Loreto, A., Di Stefano, M., Gering, M. & Conforti, L. Wallerian degeneration is executed by an NMN-SARM1-dependent late Ca2+ influx but only modestly influenced by mitochondria. Cell Rep. 13, 2539–2552 (2015).
Gerdts, J., Brace, E. J., Sasaki, Y., DiAntonio, A. & Milbrandt, J. SARM1 activation triggers axon degeneration locally via NAD+ destruction. Science 348, 453–457 (2015).
Zhao, X. F. et al. The injured sciatic nerve atlas (iSNAT), insights into the cellular and molecular basis of neural tissue degeneration and regeneration. Elife 11, e80881 (2022).
Kopp, W. How western diet and lifestyle drive the pandemic of obesity and civilization diseases. Diabetes, Metab. Syndr. Obes. 12, 2221–2236 (2019).
Clemente-Suárez, V. J., Beltrán-Velasco, A. I., Redondo-Flórez, L., Martín-Rodríguez, A. & Tornero-Aguilera, J. F. Global impacts of Western diet and its effects on metabolism and health: a narrative review. Nutrients 15, 2749 (2023).
Elafros, M. A., Reynolds, E. L. & Callaghan, B. C. Obesity-related neuropathy: the new epidemic. Curr. Opin. Neurol. 37, 467–477 (2024).
Softic, S. et al. Dietary sugars alter hepatic fatty acid oxidation via transcriptional and post-translational modifications of mitochondrial proteins. Cell Metab. 30, 735–753 (2019).
Softic, S. et al. Divergent effects of glucose and fructose on hepatic lipogenesis and insulin signaling. J. Clin. Invest. 127, 4059–4074 (2017).
Sherwani, S. I., Khan, H. A., Ekhzaimy, A., Masood, A. & Sakharkar, M. K. Significance of HbA1c test in diagnosis and prognosis of diabetic patients. Biomark Insights 11, 95–104 (2016).
Han, B. G. et al. Markers of glycemic control in the mouse: comparisons of 6-h- and overnight-fasted blood glucoses to Hb A1c. Am. J. Physiol. Endocrinol. Metab. 295, E981–986 (2008).
Themistocleous, A. C. et al. The pain in neuropathy study (PiNS): a cross-sectional observational study determining the somatosensory phenotype of painful and painless diabetic neuropathy. Pain 157, 1132–1145 (2016).
Handzlik, M. K. et al. Insulin-regulated serine and lipid metabolism drive peripheral neuropathy. Nature 614, 118–124 (2023).
O’Brien, P. D., Sakowski, S. A. & Feldman, E. L. Mouse models of diabetic neuropathy. ILAR J. 54, 259 (2014).
Gerdts, J., Summers, D. W., Sasaki, Y., DiAntonio, A. & Milbrandt, J. Sarm1-mediated axon degeneration requires both SAM and TIR interactions. J. Neurosci. 33, 13569 (2013).
Osterloh, J. M. et al. dSarm/Sarm1 is required for activation of an injury-induced axon death pathway. Science 337, 481–484 (2012).
Figley, M. D. et al. SARM1 is a metabolic sensor activated by an increased NMN/NAD+ ratio to trigger axon degeneration. Neuron 109, 1118–1136 (2021).
Berbudi, A., Rahmadika, N., Tjahjadi, A. I. & Ruslami, R. Type 2 diabetes and its impact on the immune system. Curr. Diabetes Rev. 16, 442 (2020).
Kolter, J. et al. A subset of skin macrophages contributes to the surveillance and regeneration of local nerves. Immunity 50, 1482–1497 (2019).
Jain, A. et al. Nociceptor-immune interactomes reveal insult-specific immune signatures of pain. Nat. Immunol. 25, 1296–1305 (2024).
Bruce, K. D. et al. Lipoprotein lipase is a feature of alternatively-activated microglia and may facilitate lipid uptake in the CNS during demyelination. Front. Mol. Neurosci. 11, 57 (2018).
Huey, P. U., Waugh, K. C., Etienne, J. & Eckel, R. H. Lipoprotein lipase is expressed in rat sciatic nerve and regulated in response to crush injury. J. Lipid Res. 43, 19–25 (2002).
Narciso, M. S. et al. Sciatic nerve regeneration is accelerated in galectin-3 knockout mice. Exp. Neurol. 217, 7–15 (2009).
Bray, E. R. et al. Thrombospondin-1 mediates axon regeneration in retinal ganglion cells. Neuron 103, 642–657 (2019).
Mietto, B. S. et al. Lack of galectin-3 speeds Wallerian degeneration by altering TLR and pro-inflammatory cytokine expressions in injured sciatic nerve. Eur. J. Neurosci. 37, 1682–1690 (2013).
Yu, C. et al. Microglia at sites of atrophy restrict the progression of retinal degeneration via galectin-3 and Trem2. J. Exp. Med. 221, e20231011 (2024).
Chen, P., Piao, X. & Bonaldo, P. Role of macrophages in Wallerian degeneration and axonal regeneration after peripheral nerve injury. Acta Neuropathol. 130, 605–618 (2015).
Weisberg, S. P. et al. CCR2 modulates inflammatory and metabolic effects of high-fat feeding. J. Clin. Invest. 116, 115–124 (2006).
Lefebvre, E. et al. Antifibrotic effects of the dual CCR2/CCR5 antagonist cenicriviroc in animal models of liver and kidney fibrosis. PLoS ONE 11, e0158156 (2016).
García-Revilla, J. et al. Galectin-3, a rising star in modulating microglia activation under conditions of neurodegeneration. Cell Death Dis. 13, 628 (2022).
Cheng, H. T., Dauch, J. R., Hayes, J. M., Hong, Y. & Feldman, E. L. Nerve growth factor mediates mechanical allodynia in a mouse model of type 2 diabetes. J. Neuropathol. Exp. Neurol. 68, 1229–1243 (2009).
Sierra-Silvestre, E., Somerville, M., Bisset, L. & Coppieters, M. W. Altered pain processing in patients with type 1 and 2 diabetes: systematic review and meta-analysis of pain detection thresholds and pain modulation mechanisms. BMJ Open Diabetes Res. Care 8, e001566 (2020).
Eid, S. A. et al. Single-cell RNA-seq uncovers novel metabolic functions of Schwann cells beyond myelination. J. Neurochem. 166, 367–388 (2023).
Boza-Serrano, A. et al. Galectin-3, a novel endogenous TREM2 ligand, detrimentally regulates inflammatory response in Alzheimer’s disease. Acta Neuropathol. 138, 251–273 (2019).
Mehina, E. M. F. et al. Invasion of phagocytic Galectin 3 expressing macrophages in the diabetic brain disrupts vascular repair. Sci. Adv. 7, eabg2712 (2021).
Pugliese, G., Iacobini, C., Pesce, C. M. & Menini, S. Galectin-3: an emerging all-out player in metabolic disorders and their complications. Glycobiology 25, 136–150 (2015).
Khalid, M., Petroianu, G. & Adem, A. Advanced glycation end products and diabetes mellitus: mechanisms and perspectives. Biomolecules 12, 542 (2022).
Forbes, J. M., Cooper, M. E., Oldfield, M. D. & Thomas, M. C. Role of advanced glycation end products in diabetic neuropathy. Curr. Pharm. Des. 14, 953–961 (2008).
Osonoi, S. et al. RAGE activation in macrophages and development of experimental diabetic polyneuropathy. JCI Insight 7, e160555 (2022).
Carmona, S. et al. The role of TREM2 in Alzheimer’s disease and other neurodegenerative disorders. Lancet Neurol. 17, 721–730 (2018).
Yuan, P. et al. TREM2 haplodeficiency in mice and humans impairs the microglia barrier function leading to decreased amyloid compaction and severe axonal dystrophy. Neuron 90, 724–739 (2016).
Lee, C. Y. D. et al. Elevated TREM2 gene dosage reprograms microglia responsivity and ameliorates pathological phenotypes in Alzheimer’s disease models. Neuron 97, 1032–1048 (2018).
O’Brien, P. D. et al. Integrated lipidomic and transcriptomic analyses identify altered nerve triglycerides in mouse models of prediabetes and type 2 diabetes. Dis. Model. Mech. https://doi.org/10.1242/dmm.042101 (2020).
Stratton, J. A. et al. Macrophages regulate Schwann cell maturation after nerve injury. Cell Rep. https://doi.org/10.1016/j.celrep.2018.08.004 (2018).
Chavakis, T., Alexaki, V. I. & Ferrante, A. W. Macrophage function in adipose tissue homeostasis and metabolic inflammation. Nat. Immunol. 24, 757–766 (2023).
Peng, C., Chen, J., Wu, R., Jiang, H. & Li, J. Unraveling the complex roles of macrophages in obese adipose tissue: an overview. Front. Med. https://doi.org/10.1007/S11684-023-1033-7 (2024).
Sun, J.-J. et al. Infiltration of blood-derived macrophages contributes to the development of diabetic neuropathy. J. Immunol. Res. https://doi.org/10.1155/2019/7597382 (2019).
Saika, F., Kiguchi, N., Matsuzaki, S., Kobayashi, D. & Kishioka, S. Inflammatory macrophages in the sciatic nerves facilitate neuropathic pain associated with type 2 diabetes mellitus. J. Pharmacol. Exp. Ther. 368, 535–544 (2019).
Vuckovic, S. M. et al. Up–Down Reader: an open source program for efficiently processing 50% von Frey thresholds. Front. Pharmacol. https://doi.org/10.3389/fphar.2018.00433 (2018).
Mangus, L. M., Rao, D. B. & Ebenezer, G. J. Intraepidermal nerve fiber analysis in human patients and animal models of peripheral neuropathy: a comparative review. Toxicol. Pathol. 48, 59–70 (2020).
Hafemeister, C. & Satija, R. Normalization and variance stabilization of single-cell RNA-seq data using regularized negative binomial regression. Genome Biol. 20, 296 (2019).
Kaiser, T. et al. MyelTracer: a semi-automated software for myelin g-ratio quantification. eNeuro 8, ENEURO.0558-20.2021 (2021).
Liu, Y. et al. Touch and tactile neuropathic pain sensitivity are set by corticospinal projections. Nature 561, 547–550 (2018).
Taub, D. G. et al. The secondary somatosensory cortex gates mechanical and heat sensitivity. Nat. Commun. 15, 1289 (2024).
Enders, J., Swanson, M. T., Ryals, J. & Wright, D. E. A ketogenic diet reduces mechanical allodynia and improves epidermal innervation in diabetic mice. Pain 163, 682–689 (2022).
Davis, A., Gao, R. & Navin, N. E. SCOPIT: sample size calculations for single-cell sequencing experiments. BMC Bioinf. 20, 566 (2019).
Acknowledgements
We thank D. Taub, N. Andrews, M. Kotoda and J. Hickey for assistance; S. Liberles, M. Lehtinen and R. Jackson for discussions; N. Biscola and L. Havton for the immunohistochemistry protocol; M. Scimone and the Boston Children’s Hospital Cellular Imaging Core; R. Mathieu and the Boston Children’s Hospital Flow Core; M. Ericsson and the Electron Microscopy Core at Harvard Medical School; and the Technology Center for Genomics and Bioinformatics at the University of California, Los Angeles. This work was supported by the US National Institutes of Health (grant numbers F31NS127357 to S.H. and R35NS105076 to C.J.W.) and the Dr. Miriam and Sheldon G. Adelson Medical Research Foundation (C.J.W. and R.K.).
Author information
Authors and Affiliations
Contributions
S.H. and C.J.W. conceptualized, and led the design and execution of the study. S.H. performed behavioural assays with help from A.J., V.P., J.I. and E.S.D. S.H. performed flow cytometry. S.S.A. performed glucose tolerance tests. S.H. performed single-cell sequencing and analysis with help from A.J., J.I., R.K. and Q.W. S.H., J.I., E.S.D. and D.N. performed immunohistochemical and IENFD analysis. A.J. and V.P. performed mouse injections. S.H. and H.W.K. performed sciatic nerve surgeries. S.H., J.I., E.S.D., D.N., C.A.G. and J.R. performed feeding experiments and weight monitoring. S.H., A.J. and C.J.W. guided experimental design and analysis. S.H. and C.J.W. wrote the manuscript with input from all authors.
Corresponding author
Ethics declarations
Competing interests
C.J.W. is a founder of Nocion, Quralis and Blackbox Bio. The other authors declare no competing interests.
Peer review
Peer review information
Nature thanks David Bennet and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data figures and tables
Extended Data Fig. 1 High Fat High Fructose feeding leads to obesity and pre-diabetes.
a. Body weights of male C57Bl6 mice fed a HFHFD or CD over six months (n = 8 mice per group, data are presented as mean values +/- SEM, p-value from 2way ANOVA). b. Glucose tolerance test after fasting in male mice fed a HFHFD or CD for 12 weeks. (n = 5 mice per group, data are presented as mean values +/- SEM, p-value from 2way ANOVA). c. Area under the curve from a glucose tolerance test at 12 weeks (n = 5 mice per group, data are presented as mean values +/- SEM, p-value from two-tailed unpaired student’s t-test). d. Fasting plasma insulin measurements from male mice fed a HFHFD or CD for 12 weeks (n = 5 mice in CD and 4 mice in HFHFD, data are presented as mean values +/- SEM, p-value from two-tailed unpaired student’s t-test). e. Percent glycated HbA1c measurements from whole blood of male mice fed a HFHFD or CD for 12 weeks (n = 5 mice per group, data are presented as mean values +/- SEM, p-value from two-tailed unpaired student’s t-test). f. Glucose tolerance test after fasting in male mice fed a HFHFD or CD for 24 weeks (n = 5 mice per group, data are presented as mean values +/- SEM, p-value from 2way ANOVA). g. Area under the curve from glucose tolerance test at 24 weeks (n = 5 mice per group, data are presented as mean values +/- SEM, p-value from two-tailed unpaired student’s t-test). h. Fasting insulin measurement from plasma of male mice fed HFHFD or CD for 24 weeks (n = 5 mice per group, data are presented as mean values +/- SEM, p-value from two-tailed unpaired student’s t-test). i. Percent glycated HbA1c measurement from whole blood of male mice fed HFHFD or CD for 24 weeks (n = 5 mice per group, data are presented as mean values +/- SEM, p-value from two-tailed unpaired student’s t-test).
Extended Data Fig. 2 Female mice on a HFHFD are equally susceptible to weight gain, behavioral changes, and immune cell infiltration in nerves.
a. Body weight measurements over time on a HFHFD vs CD from female C57Bl6 mice. (n = 8 mice per group, data are presented as mean values +/- SEM, p-value < 0.0001 from 2way ANOVA) b. Von Frey up down method was used to assess mechanical sensitivity in female mice fed a HFHFD vs CD over 12 weeks. (n = 8 mice per group, data are presented as mean values +/- SEM, p-values from 2way ANOVA with Sidak multiple comparisons) c. Hargreaves assay was used to assess heat sensitivity in female mice fed HFHFD vs CD over 12 weeks. (n = 8 mice per group, data are presented as mean values +/- SEM, p-values from 2way ANOVA with Sidak multiple comparisons) d. Proportions of Ly6c+ CD64+ recruited macrophages from Live Cd45+ Cd11b+ cells in sciatic nerves of female mice fed a HFHFD vs CD for 12 weeks. (n = 8 mice per group, data are presented as mean values +/- SEM, p-value from two-tailed unpaired student’s t-test).
Extended Data Fig. 3 Sarm1 knockdown prevents the loss of heat sensitivity and skin axonal degeneration in HFHFD-fed mice but leads to persistent mechanical allodynia.
a. Illustration of the experiment. b. Body weight of WT and Sarm1-KO mice over time on a CD or HFHFD. (n = 5 mice per group, data are presented as mean values +/- SEM). c. Thermal sensitivity in WT vs Sarm1-KO male mice fed a HFHFD or CD over 24 weeks. (n = 5 mice per group, data are presented as mean values +/- SEM, p-values from 2way ANOVA with Sidak multiple comparisons). d. Mechanical sensitivity in WT vs Sarm1-KO male mice fed a HFHFD or CD over 24 weeks. (n = 5 mice per group, data are presented as mean values +/- SEM, p-values from 2way ANOVA with Sidak multiple comparisons). e. Quantification of IENFD from PGP9.5 staining in hind paw skin from WT and Sarm1-KO male mice fed a HFHFD or CD for 24 weeks. (n = 5 mice per group, data are presented as mean values +/- SEM, p-value from two-tailed unpaired student’s t-test). f. Summary of findings from Sarm1-KO mouse strain compared to WT mice on a HFHFD. The graphics in a were created with BioRender.com.
Extended Data Fig. 4 Sciatic nerve immune cell single cell sequencing.
a. Illustration of experimental workflow. b. Number of cells analyzed in each sample post-filtering. c. Dot plot of gene markers used to identify cell types in each cluster. d. UMAP plot showing similar cell composition in replicates of each condition. The graphics in a were created with BioRender.com.
Extended Data Fig. 5 Gating strategy for a flow cytometry assessment of recruited macrophages into the sciatic nerve.
a. FSC-A and SSC-A were used to gate cells and exclude aggregates and small debris. b. FSC-A and FSC-W were used to gate single cells. c. DAPI and CD45-APC-ef780 were used to capture live CD45+ immune cells. d. Cd11b-FITC and Ly6g-PE were used to gate myeloid cells and exclude neutrophils. e. Cd64-PE-594 and Ly6c-BV711 were used to identify Ly6C+ Cd64+ recruited macrophages. (n = 5 mice per group).
Extended Data Fig. 6 Relationship of recruited macrophages to vasculature and identification of source of CCL2.
a. Representative cross section image from a sciatic nerve from mice fed a CD or HFHFD for 12 weeks staining for Cd31 (vasculature), F4/80 (macrophages), and DAPI (nuclei). b. Quantification of RecMacs (Cd64+Ly6c+) macrophages within the live Cd45+Cd11b+Ly6g- myeloid cell population following a perfusion with PBS in mice fed a CD or HFHFD for 12 weeks. (n = 4 mice in CD and 5 mice in HFHFD group, data are presented as mean values +/- SEM, p-value from two-tailed unpaired student’s t-test). c. Representative image from sciatic nerves of Ccl2-RFP mice fed a CD or HFHFD for 12 weeks showing no overlap with PGP9.5 (neurons) (representative from 2 independent experiments, n = 3 mice per group each, scale bar = 100 um). d. Representative image from sciatic nerves of Ccl2-RFP mice fed a CD or HFHFD for 12 weeks showing no overlap with S100 (Schwann Cells) (representative from 2 independent experiments, n = 3 mice per group each, scale bar = 100 um).
Extended Data Fig. 7 HFHFD feeding does not lead to demyelination or evidence of myelin disturbances after 12 weeks in the sciatic nerve.
a. Representative EM images from sciatic nerves of mice fed a CD or HFHFD for 12 weeks. b. Scatterplot and slope of each quantified axon diameter vs. g-ratio from CD samples and HFHFD samples. (n = 3 mice on CD and 4 mice on HFHFD, data are presented as mean values +/- SEM). c. Quantification of average axon diameter per mouse from 3 non-consecutive images. (n = 3 mice on CD and 4 mice on HFHFD, data are presented as mean values +/- SEM, p-value from two-tailed unpaired student’s t-test). d. Quantification of g-ratio split by axon diameter ranges per mouse from 3 non-consecutive images. (n = 3 mice on CD and 4 mice on HFHFD, data are presented as mean values +/- SEM, p-values from multiple unpaired t-tests corrected using Two-stage step-up method (Benjamini, Krieger, Yekutieli)). e. Quantification of percent myelinated fibers per mouse from 3 non-consecutive images. (n = 3 mice on CD and 4 mice on HFHFD, data are presented as mean values +/- SEM, p-value from two-tailed unpaired student’s t-test). f. Quantification of average number of axons per Remak bundle per mouse from 3 non-consecutive images. (n = 3 mice on CD and 4 mice on HFHFD, data are presented as mean values +/- SEM, p-value from two-tailed unpaired student’s t-test).
Extended Data Fig. 8 Analysis of skin and dorsal root ganglia immune cells from mice fed a HFHFD or CD for 12 weeks shows no change in immune cell composition.
a. Workflow for tissue collection, processing, sorting, and sequencing of skin immune cells from male mice fed a HFHFD or CD for 12 weeks with 4 mice per group pooled into 2 samples. b. Number of cells analyzed in each sample post-filtering. c. Dot plot of marker genes used to identify different immune cells in dataset. d. UMAP plot showing clusters identified in sciatic nerves of CD and HFHFD fed mice from 2 samples per group. e. Barplot showing proportions of the different cell types per group. f. Flow cytometry analysis of recruited macrophages (Ly6c+Cd64+) within the myeloid cell population (Live Cd45+Cd11b+Ly6g-). (n = 3 mice in CD ad 5 mice in HFHFD group, data are presented as mean values +/- SEM, p-value from two-tailed unpaired student’s t-test). g. Volcano plot of differentially expressed genes in skin resident macrophages in HFHFD samples compared to CD, dashed lines at p-value = 0.05 and Log2- Fold Change −0.5 and 0.5. h. GO molecular function analysis of upregulated genes in resident macrophages. i. Representative flow cytometry contour plots from DRG immune cells from CD and HFHFD-fed mice for 12 weeks pre-gated on live Cd45+ Cd11b+ Ly6g- cells showing Ly6c and Cd64 staining. j. Flow cytometry quantification of recruited macrophages (Ly6c+Cd64+) within the myeloid cell population (Live Cd45+Cd11b+Ly6g-). (n = 5 mice per group, data are presented as mean values +/- SEM, p-value from two-tailed unpaired student’s t-test). The graphics in a were created with BioRender.com.
Extended Data Fig. 9 Mice lacking CCR2-KO show no overt difference in glycemic status, T cells, or mechanical thresholds.
a. Body weights from male WT and Ccr2-KO mice fed a HFHFD over time (n = 5 mice per group, data are presented as mean values +/- SEM). b. Glucose Tolerance Test from male WT and Ccr2-KO mice fed a HFHFD for 12 weeks (n = 5 mice per group, data are presented as mean values +/- SEM) c. Area under the curve from glucose tolerance test from male WT and Ccr2-KO mice fed a HFHFD for 12 weeks (n = 5 mice per group, data are presented as mean values +/- SEM, p-value from two-tailed unpaired student’s t-test). d. Percent glycated HbA1c from whole blood from male WT and Ccr2-KO mice fed a HFHFD for 12 weeks (n = 5 mice per group, data are presented as mean values +/- SEM, p-value from two-tailed unpaired student’s t-test). e. Fasting insulin levels measured from plasma of male WT and Ccr2-KO mice fed a HFHFD for 12 weeks (n = 5 mice per group, data are presented as mean values +/- SEM, p-value from two-tailed unpaired student’s t-test). f. Flow cytometry analysis of T Cells (Cd3+) within live Cd45+ immune cells (n = 5 mice per group, data are presented as mean values +/- SEM, p-value from two-tailed unpaired student’s t-test). g. Mechanical sensitivity assessed using a von Frey assay in male WT and Ccr2-KO mice fed a HFHFD (n = 5 mice per group, data are presented as mean values +/- SEM, p-values from 2way ANOVA with Sidak multiple comparisons). h. Heat sensitivity assessed using Hargreaves assay in male WT and Ccr2-KO mice fed HFHFD for 24 weeks (n = 5 mice per group, data are presented as mean values +/- SEM, p-value from two-tailed unpaired student’s t-test).
Extended Data Fig. 10 CCR2 blockade has no effect on CD fed mice, leads to increased warm preference after HFHFD and the effect of Lgals3 knockdown is not mediated by reduced recruitment of macrophages to the nerve.
a. Body weights from male WT mice fed a HFHFD over time injected with CVC or Vehicle (n = 4 mice per group, data are presented as mean values +/- SEM). b. Mechanical sensitivity was assessed using the von Frey up down method in mice at 8 weeks on a HFHFD and at 12 weeks following CVC or Veh injections (n = 9 CD and 8 HFHFD, data are presented as mean values +/- SEM). c. Heat sensitivity assessed using Hargreaves assay in mice fed a CD for 12 weeks and injected with CVC or Veh starting after 8 weeks on diet. (n = 7 mice per group, data are presented as mean values +/- SEM). d. quantification of IENFD from mice fed a CD for 12 weeks and injected with CVC or Veh starting after 8 weeks on diet. (n = 7 mice per group, data are presented as mean values +/- SEM). e. Time spent per zone on a thermal gradient ring (n = 5 WT and n = 4 KO female mice, data are presented as mean values +/- SEM) (p-value at 28 °C < 0.0001, p-value at 10 °C = 0.1963, 14 °C = 0.2177, 17 °C = 0.1284, 21 °C = 0.4804, 24 °C = 0.8515, 28 °C < 0.0001, 31 °C = 0.1923, 35 °C = 0.9993, all other p-values >0.9999 from 2way ANOVA with Sidak multiple comparisons). f. Time spent in different temperature zones on thermal gradient ring (n = 4 male mice per group, data are presented as mean values +/- SEM) (p-value at 28 °C = 0.5052, 31 °C = 0.0105, 35 °C = 0.4037, at 39 °C = 0.0135, all other p-values >0.9999 from 2way ANOVA with Sidak multiple comparisons). g. Body weight at start and end of experiment after HFHFD feeding (n = 5 WT and n = 4 KO, data are presented as mean values +/- SEM). h. Quantification of recruited macrophages (recMacs) (Ly6c+Cd64+) from nerves within the live Cd45+Cd11b+ Ly6g- population using flow cytometry or WT or Lgals3-KO mice fed HFHFD for 12 weeks (n = 7 mice per group, data are presented as mean values +/- SEM, p-value from two-tailed unpaired student’s t-test).i. Illustrative summary of the findings described in this manuscript created with BioRender.com.
Supplementary information
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Hakim, S., Jain, A., Adamson, S.S. et al. Macrophages protect against sensory axon loss in peripheral neuropathy. Nature 640, 212–220 (2025). https://doi.org/10.1038/s41586-024-08535-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41586-024-08535-1
