
Abstract
The continuous exposure of chemical pesticides in agriculture, their contamination in soil and water pose serious threat to the environment. Current study used an approach to evaluate various pesticides like Hexaconazole, Mancozeb, Pretilachlor, Organophosphate and λ-cyhalothrin degradation capability of esterase. The enzyme was isolated from Salinicoccus roseus. Genome analysis unveiled the carboxylesterase genes underlying the degradation of pesticides, and was located between 2070Mbp to 2080Mbp region. Herein, partially purified esterase was immobilized into beads by mixing with an equal volume (1:1) of sodium alginate solution [2.5% (w/v)].Scanning electron microscopy (SEM) of the beads showed the microspheres for enhanced enzyme-substrate reaction, wide peak at 3316, 1635 and 696 cm− 1 in Fourier-transform infrared spectroscopy (FTIR) represented intermolecular hydrogen bonding, and thermogravimetric analysis (TGA) reaffirmed the binding of esterase entrapped into the beads. Maximum degradation rate (after 4 days) for free enzyme accounted 83.2% in Hexaconazole. Degradation rate moderately increased 4% in the presence of immobilized esterase. Degradation products were detected by liquid chromatography–mass spectrometry (LC–MS). Cytotoxicity test (root length and mitotic index) revealed differences in various treatments. Enzyme kinetics parameters, Michaëlis-Menten constant (KM) 6.61 mM and maximum velocity (Vmax) 1.89 µmol/min/mg increased after immobilization. Further, molecular docking results validated that esterase contributed to pesticide degradation by catalytic triad of Ser93-His222-Phe24, ligand interactions, and specific binding pockets. Additionally, molecular dynamics (MD) simulations confirmed the protein-ligand conformational stability. Hence, present study highlighted an effective method for improving the catalytic properties of esterase, and also potential candidate for bioremediation of pesticides.
Similar content being viewed by others

Introduction
Pesticide contamination is one of the rising challenges of today’s world because of their biomagnification and persistent nature1. Some of the pesticides such as organophosphates (OPs) organochlorines (OCs) are dangerous chemicals and remained in the environment as persistent organic pollutants (POPs). Pesticides are a group of chemical substances used in agricultural lands that incapacitate pests and weeds2. The World Health Organization (WHO) estimated over 150,000 deaths occurs annually by accidental or intentional exposure to pesticides3. Pesticide residues are able to reach different compartments of the ecosystem through a repeated process of evaporation and deposition, consequently, evidence is mounting that agricultural lands are affected significantly4. Concurrently, pesticide effects on non-target organisms have been a source of worldwide concern. For example, presence of POPs residues in human breast milk, raising worries about fetal exposure and adverse health effects on infants5. Therefore, the residual concentration present in the soil must be reduced. The relatively new concept of pesticide bioremediation provides more greener, economic, and environmentally competent alternative than the traditional physical and chemical methods6.
Depending on the target pests, different pesticide applications can be categorized as fungicides, herbicides, or insecticides etc7. Based on the chemical composition and active ingredients, pesticides were classified into several groups like Hexaconazole, Mancozeb, Pretilachlor, Organophosphate and λ-cyhalothrin. Hexaconazole is an active broad-spectrum 1,2,4-triazole fungicide with a pair of enantiomers, displaying higher fungicidal activity [(-)-form] than the other enantiomer [(+)- form]8. Mancozeb is a non-systemic dithiocarbamates contact fungicide9. Pretilachlor is one of the well-known synthetic herbicides, and belongs to the chloroacetamide group10. Organophosphate pesticides were developed in the 1950s and account for about 40% of the total pesticides around the world11. The chiral insecticide, λ-cyhalothrin possesses eight stereoisomers and exhibits greater risk to non-target organisms12.
Esterases (EC 3.1.1.X) are catalysts that hydrolyze water soluble or emulsified esters and synthesize ester bonds across a large variety of substrates. Carboxylesterase is one of the widely used enzymes applied in the resolution of racemic drugs from a mixture of chiral drugs and pesticides13. Previous reports mentioned that esterase played a noteworthy role in detoxification and effective biodegradation of chemical pesticides14. The degradation of pesticides such as Hexaconazole through the catalytic activity of esterase, isolated from Salinicoccus roseus strain RF1H has been facilitated in this study. The problem with free enzymes is that often undergo dissociation and had negligible reusability. In order to improve enzyme catalytic properties, immobilization showed remarkable stability and can efficiently remove biological pollutants15.The encapsulation material chosen to immobilize esterase was natural biopolymer alginate formed by the glycosidic bond between β-D-mannuronic acid and α-L-guluronic acid16. Besides its non-toxicity, low cost and availability, sodium alginate is deciphered with excellent mechanical strength and flexibility17.
Although, pesticide degradation is well understood through enzyme encapsulated beads, we attempted to extend our knowledge of bioremediation using a bioinformatics approach. The molecular docking tool provides insight into enzyme-substrate interaction, whether the active site accommodates the pollutant or not18. We pointed here esterase-pesticide binding characteristics by molecular docking analysis. Furthermore, molecular dynamics (MD) simulation was executed to clarify the dynamic behavior and the docking results as well.
For the purpose of the production of various enzymes, numerous bacteria are frequently used in biotechnology and industrial processes19. Fusion protein technology enables Clostridium sp. to improve enzyme production yield and organophosphate degradation20. Genes encoding pesticides degrading esterase like CesB in Bacillus subtilis has been documented by many studies21. The analysis of genome data from S. roseus may improve our understanding of the carboxylesterase gene and biodegradation of different hydrocarbons by the strain. Therefore, the present study can provide an insight into the biochemical and genetic outlook of esterase and hexaconazole compound.
To address the above problems of pesticide residues in the environment and dissociation of free enzymes, this study was carried out with following objectives- (i) to prepare enzyme for encapsulation (ii) to characterize the biophysical nature of encapsulated enzyme (iii) to apply the immobilized enzyme in degradation of pesticides (iv) to analyze most potent degradation products and investigate their toxicity (v) to determine the enzyme kinetics for binding affinities of substrates and molecular docking for active site prediction (vi) to find out genomic determinants responsible for pesticide degradation in S. roseus.
Results
Genome analysis of RF1H
The draft whole genome sequence data of Salinicoccus roseus strain RF1H has been submitted to NCBI GenBank under the Accession number JAIMFU010000000.1; Bio project number PRJNA756885. Different hydrocarbon constituents of various pesticides like benzoate, salicylate ester and anthracene degradation clusters were found in the genome (Fig. 1a) (Table S1). AreA for aryl ester degradation; benzoate MFS transporter BenK; Naphthalene 1,2-dioxygenase system ferredoxin; 3-phenylpropionate dioxygenase genes were reported from the genome. In addition, a comparative genomic analysis of six S. roseus genomes available in NCBI GenBank (DSM 5351, W12, MOSEL-ME25, CCM 3516, BU-1, and RF1H) (accessed on May 17, 2022) confirmed that the carboxylesterase gene was acquired in 2070Mbp to 2080Mbp region of RF1H genome (Fig. 1b). No occurrence of carboxylesterase gene within the total 17 genomic islands (GIs) strongly suggested that these genes have been horizontally acquired. This could be an adaptive mechanism to different environmental stresses. Thus, combining the wet lab data and genome information of S. roseus deciphered the role of carboxylesterase in pesticide degradation.

Genomic information of Salinicoccus roseus strain RF1H (a) Whole genome annotation (subsystems and categories were classified using RAST server) (b) genome comparison between six S. roseus strains, showing shared region for hydrocarbon degrading esterase gene cluster (circular map was created using CGView server).
Partial purification of esterase
Crude esterase precipitation was recorded at 70% ammonium sulfate precipitation. A purification fold of 3.37 with a specific activity of 5.06 U/mg was determined (Table 1).
Physico-biochemical properties of the beads
The external diameter of the alginate bead was 2.0 mm. Surface morphology of the esterase immobilized particles showed the formation of pores, small particles, and microspheres which had created a microenvironment for enhanced enzyme-substrate reaction. The surface of bare alginate beads appeared smooth (Fig. 2a) whereas the rough surface of the esterase-loaded beads clearly indicated that the gel had restricted esterase into the matrix (Fig. 2b).

Scanning electron micrographs of sodium alginate beads (a) without esterase (b) immobilized esterase. The micrographs displayed surface structure of alginate beads, whereas enzyme loaded beads had a rough surface appearance.
The FTIR spectrum of enzyme loaded beads has been shown in Fig. 3a. The significant wide peak at 3316 cm− 1 represented the hydroxyl groups, including stretching vibrations of N-H and O-H bonds present in the sodium alginate polymer chain. The next peak appeared at 1635 cm− 1, attributed to the bending vibration of N–H. The absorption peak of esterase showed at 1990 –1950 cm− 1 corresponded with C = O symmetric stretching. Then the peak around 1240 –1195 cm− 1 characterized the deformation of the amide N-H group and those at 1013 cm− 1 could be assigned to symmetric and asymmetric vibration bands of C-O-C bonds, which were present in polysaccharide rings. This result confirmed the interaction between sodium alginate and esterase took place successfully. The diffractogram of sodium alginate bead has been represented in Fig. 3b. Two major crystalline peaks at 2θ diffraction angle of 30 and 42, indicated the semi-crystalline nature of alginate. The thermogram profile of immobilized esterase in Fig. 3c showed a weight loss at 170 °C, likely to occur due to dehydration. Another significant weight loss was observed at higher temperatures for the heating rate of 260 °C and 340 °C, corresponding to the decomposition of glycosidic bonds of alginate followed by sample degradation process respectively.

Characterization of sodium alginate immobilized esterase beads (a) FTIR spectrum of Na-alginate-esterase beads showing different bands corresponding to alginate and esterase interaction in the spectrum scale of 4000–400 cm− 1 (b) X-ray diffractogram of the Na-alginate-esterase beads representing crystalline nature of alginate (c) Thermogravimetric curve of Na-alginate heated at 10 °C/min where a weight loss at 170 °C was identified.
Application of esterase for pesticides degradation
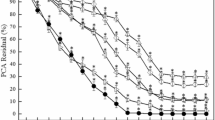
Among all the tested pesticides, Hexaconazole showed maximum degradation percentage by the application of free esterase as well as immobilized enzyme (Fig. 4 and Table S2). It had been proved that immobilized esterase could effectively catalyze the pesticides. An increment of 4% in degradation rate was observed in Hexaconazole, compared to the application of free enzyme. Analysis with two-way ANOVA also revealed significant difference between these two applications (p < 0.05).

Esterase assisted degradation of various pesticides.
Metabolites identification of pesticide degradation
The metabolites for Hexaconazole degradation were detected by LC-MS analysis. The products identified by mass spectrometry analysis were matched with Chemistry WebBook data of National Institute of Standards and Technology (NIST, USA) and PubChem (NIH, USA). Apart from the parent compound Hexaconazole, retention time (RT), 1.93 min; quantitation ion (m/z): 217.10, m/z: 261.13, m/z: 305.15 products were found (Fig. 5a). Three sharp peaks, shown in Fig. 5b, (RT 13.94 min) corresponded to the compounds- (m/z: 302.30) 1-(2,4-dichlorophenyl)-2-(1 H-1,2,4-triazol-1-yl) ethenone; (m/z: 318.30) 2-(2, 4-dichlorophenyl)-1-(1 H-1,2,4-triazol-1-yl) hexane-2; 5-diol, and (m/z: 369.35) 1-(2, 4-dichlorophenyl)-2-(1 H-1,2,4-triazol-1-yl) ethanol. The disappearance of parent and origin of other peaks suggested that esterase followed biodegradation pathway or further biodegraded into smaller compounds.

LC-MS analysis of the metabolites produced after the application of esterase (a) at retention time 1.93 min (b) at retention time 13.94 min [generated compounds: (i) 1-(2,4-dichlorophenyl)-2-(1 H-1,2,4-triazol-1-yl) ethenone (ii) 2-(2, 4-dichlorophenyl)-1-(1 H-1,2,4-triazol-1-yl) hexane-2; 5-diol (iii) 1-(2, 4-dichlorophenyl)-2-(1 H-1,2,4-triazol-1-yl) ethanol]
Toxicity studies
The root lengths of Allium cepa after exposure to different times showed significant variations (Fig. S1). The maximum root length was observed in control (distilled water) with 72 h exposure. Inhibition of root growth by Hexaconazole treatment is an indicator of growth retardation of toxicity. The effect of cytotoxicity was observed under microscope.
Normal mitotic stages of root meristem cells were observed in control (Fig. 6a-d).Different type of chromosomal aberrations were noted as anaphase bridge, C-mitosis and fragments (Fig. 6e–g). According to the results in Table 2, MI decreased significantly in Hexaconazole treated root tip cells at each exposure time, compared to control. In addition, all MI results were found statistically significant (p < 0.05).The MI in the root tip cells, treated with the pesticide degraded solution was higher than the pesticide itself. This was a clear indication that esterase treated solution played significant role in promotion of cell division.

Root tip cells of Allium cepa showing normal stages of mitosis [(a) Prophase (b) Metaphase (c) Anaphase (d) Telophase] and anomalies induced by Hexaconazole treatment [(e) Anaphase bridge (f) C-mitosis (g) Fragments][Scale bars 10 μm].
Enzyme kinetics
KM value and Vmax value of free esterase in presence of the substrate Hexaconazole were 5.96 mM and 1.73 µmol/min/mg respectively (Fig. 7a). However, the KM value increased after entrapment into the beads with a calculated value of 6.61 mM, and the Vmax value of 1.89 µmol/min/mg (Fig. 7b). Increased KM value in immobilized esterase suggested low availability of substrate for binding into the active site. kcat value of free esterase and immobilized esterase were 0.057s− 1 and 0.063s− 1 respectively.

Kinetic behavior of esterase using Lineweaver–Burk plot (a) before immobilization (b) after immobilization. The enzyme activity measured at various concentrations of para-Nitrophenyl acetate at 37 °C temperature. Where “S” denoted substrate concentrations and “V” denoted enzyme-specific activity.
Molecular docking analysis
The constructed model was subjected to validation using various servers. The Ramachandran plot for the predicted model had 91.1% residues in the most favoured regions, suggested a good quality model. The Verify 3D server predicted that 86.53% of the amino acids residues had an average 3D-1D score ˃=0.1, Z-score was within the range. We finally verified the structure using the ERRAT graph. ERRAT was a so-called “overall quality factor”, which analysed the statistics of nonbonded interactions between different atom types. ERRAT score of 98.31% confirmed the overall model quality (the accepted limit was > 50 for a high quality model). Verification of the results invariably indicated a good quality of the proposed model and reliability to perform further structural analysis (Table S3).
Docking studies showed that the ligand had compatible binding energies with a modelled enzyme. The highest binding energy of pesticide was found in Hexaconazole with − 5.84 kcal/mol (Table 3). MD simulation was carried out to calculate the RMSD of protein-ligand complexes for validation and confirmation. The low average RMSD values observed in the five protein-ligand complexes (Fig. 8a) revealed that the ligands and proteins were closely linked. Taking consideration for binding free energy and less fluctuating RMSD after 1 ns simulation, esterase complexed with Hexaconazole, considered as most stable.

Protein-ligand docking in the context of pesticides (a) MD simulation of esterase-pesticide complexes at 300 K using the explicit solvent model. The RMSD of the ligand and protein calculated from MD simulation of (i) Hexaconazole (ii) Mancozeb (iii) Pretilachlor (iv) Organophosphate (v) λ-cyhalothrin with esterase had been illustrated in blue, purple, pink, aqua, and green colours. The simulation not only reached to the known crystal–structure complex, which was the most thermodynamically stable state, but it also got to the unknown crystal–structure complex, which was the most thermodynamically stable state (b) RMSD graph of protein-ligand complex during 100 ns MD simulation.
We observed a decrease in the overall G -factor after MD simulation, which indicated that there might be an increase in the number of the bad dihedral angles of the modeled structure, which might be possibly due to the MD simulation resulting in an unfavorable dihedral angle, allowing the protein to overcome high-energy barriers. Further, the best-docking scored complex (Hexaconazole-esterase) was chosen to check the stability of structure during 100 nanoseconds (ns) simulation period. The results obtained during MD simulation were more significant and stable than the docked conformation. The visual analysis of the trajectory confirmed the stability of the protein at least in the time period of 100 ns (Fig. 8b).Fig. S2 confirmed that the substrates were fitted well in the active site and provided a stable environment for the catalytic residues. The amino acids formed catalytic triad with Ser93His222Phe24 in all the cases (Table S4), which hydrolyze the ester/amide/carbamate bond in the pesticides. The reaction employed in Fig. S3, serine residue by nucleophilic attack transfer nitrogen atom to the scissile bond of histidine that leading to the formation of the first tetrahedral intermediate. The intermediate broken down to acyl-enzyme complex. Acylation was the process of adding acyl group by Friedel-Craft reaction22. As a result of the acylation step, the proton of serine residue transferred to the imidazole group in the side chain of histidine. This shifting of the carbonyl atom made it prone to another nucleophilic attack by a water molecule. Thus, a second tetrahedral intermediate was formed, followed by the release of alcohol and carboxylic acid. The target-ligand interaction in molecular docking appeared to be mainly a strong hydrophobic bond. The most common were conventional hydrogen bond in all the compounds, whereas no van der Waals bonding was observed in any cases (Fig. 9a i-v). A comprehensive depiction of the 3D interactions between Hexaconazole and the active site residues of esterase have been visually presented in Fig. 9b.

Docking pose and binding interaction diagram (a) Two dimensional structures of the esterase-pesticides complexes [(i) Hexaconazole (ii) Mancozeb (iii) Pretilachlor (iv) Organophosphate (v) λ-cyhalothrin] (b) Three dimensional structure of the esterase-hexaconazole complex.
Discussion
In this study, alginate derived gel beads were prepared by using the immobilization technique. Sodium alginate has been considered because of its high economic value and its non-toxic physiological nature. The extracellular esterase from Salinicoccus roseus culture supernatant was precipitated by adding ammonium sulphate. The maximum esterase activity was achieved at 80% ammonium sulphate saturation. In contrary to our result, esterase from fungi-derived Aureobasidium pullulans was found to have best separation with 80% saturation of (NH4)2SO423. In other study, esterase from halotolerant Salimicrobium sp. showed 60% saturation in (NH4)2SO4 with 2.3-fold purification24. He et al. reported immobilized Feruloyl esterase on magnetic nanoparticles (FAE-MNPs), achieved maximum activity 10.2 ± 0.22 U/g MNPs25. However, MNPs were prone to oxidation under ambient conditions and were easily integrated with an acid medium26. Tercan et al. obtained an immobilization yield of 69.5% for the recombinant esterase of Geobacillus sp. on chitosan-alginate blended beads27.The immobilization yield of 78% for carboxylesterase was observed at 2% (w/v) sodium alginate28. In our experiment, 2.5% (w/v) sodium alginate provided the best results. The average size of the immobilized beads was approximately 2.0 nm in diameter with spherical morphology. Raghu & Pennathur observed ~ 1.6 μm sized beads, produced from Aeromonas caviae esterase showed the best activity28, whereas recombinant esterase had maximum activity at 0.5 mm diameter silicate-coated Ca-alginate beads29. This could be due to the fact that smaller sized beads reduced surface transfer resistance resulting in higher catalytic activity. XRD spectrum demonstrated that structural change of alginate occurred from grain to crystalline state but no change in the enzyme structure. Similar to our results, micro-crystallite nature in the amorphous structure of sodium alginate with characteristics peak at 35° was reported16. The results indicated that this approach was a promising tool for protecting enzymes from dissociation without hampering catalytic activity. The mass losses obtained by thermogravimetry were detected in three regions. Regions I and II (170 °C and 260 °C) were linked with the desorption of physically adsorbed water and decomposition of the organic compound of alginate beads. Soares et al. observed a similar behaviour in sodium alginate and explained the onset of weight loss due to thermal degradation30. This thermal decomposition behavior of polyanionic sodium alginate and cationic proteins of esterase explained the electrostatic interaction between them31. FTIR analysis confirmed that the immobilized enzyme has functional groups including hydroxyl, carboxyl, and amide groups that served as reducing agents to stabilize the interaction. Our result was in agreement with another available result32.
Although many organophosphate hydrolyzing enzymes have been identified and characterized22,33, halophilic carboxylesterase is the first reported enzyme with broad-spectrum pesticide hydrolyzing activity from S. roseus strain RF1H. Carboxylesterase could be potential degrading enzyme because they split the ester bonds from organophosphates, carbamates and pyrethroids, and yielded acid and alcohol34.This corroborated the findings of Xiao et al. where pyrethroid degradation efficacy of carboxylesterase from Bacillus subtilis was reported21. In research explored by Singh et al., around 36% malathion organophosphate was degraded by carboxylesterase for 7 days of incubation at 30 °C35. Here, partially purified esterase from S. roseus could catalyze organophosphate pesticides (82.9%) at acidic pH (pH 5.0), at 37 °C temperature for 4 days incubation. LC-MS results confirmed that Hexaconazole could be degraded by the catalytic activity of esterase.
In A. cepa root growth inhibition was related to inhibition of cell elongation during differentiation. Similar growth retardation was reported by Mercado and Caleño in Glyphosate herbicide toxicity36. Reduction of MI and frequent chromosomal aberrations in Hexaconazole treated meristems might be due to mito-depressive action of the compound, that interfered with normal cell cycle. Indirect genotoxic effect was recorded in diclofop-methyl herbicide37.The calculated KM for esterase in the presence of p-NPA was 9.98mM, and in the presence of Hexaconazole it was 5.96 mM38. Decreasing KM under Hexaconazole stress condition indicated that, the competition was lower than normal. In this study, KM increased to 6.61 mM after immobilization, indicating a higher concentration of substrate (Hexaconazole) was needed for the enzyme. An increase in KM value designated apparently reduced esterase affinity towards substrate, Hexaconazole. Some factors could affect the enzyme affinity like, conformational changes of enzyme when adsorbed into the beads attributed to steric hindrance of the active site39. Similar phenomenon was reported in literature where the immobilized enzyme exhibited a higher KM value than the free enzyme40.Contrary, Gülay & Şanlı-Mohamed explored lower KM value after immobilization than the free esterase12. The Vmax values of free and immobilized thermoalkalophilic recombinant esterase were 19.02 and 12.85 mmol L− 1 mL-1min− 1, respectively27. A decrease in the Vmax value of immobilized enzymes was a common phenomenon because of limited diffusion. Also, as a result of diffusional limitation and/or conformational changes of the enzyme resulting in a lower affinity of the substrate thus increased the KM39. In contrast, an increase in both Vmax and KM was noted in our result. KM value was higher in S. roseus esterase compared with the pyrethroid-hydrolyzing carboxylesterase EstSt7, which reflected the diffusional effect of the alginate matrix41. Karakuş et al. noted that Vmax value increased after immobilization, while KM value was roughly equivalent to that of the free and immobilized pectinesterase42. This could be interpreted as the loss of entropy on the binding of enzyme-substrate complex which offered catalytic advantage by intramolecular reactions43. This suggested that the structural conformation of the enzyme remains intact after immobilization. kcat value reflected the turnover number and enzyme efficiency in catalyzing reactions. The increase in turnover number for the immobilized esterase indicated higher catalytic activity of the enzyme. The kcat value of free esterase was 0.057s− 1 in hexaconazole, whereas pyrethroid-hydrolyzing carboxylesterases (PytH) recorded 3.03 ± 0.15s− 1 for permethrin44.
Molecular docking results confirmed the active binding sites of Hexaconazole. The amino acids, Ser-His-Phe formed a catalytic triad which played a major role in the catalysis of the pesticides. Compared with thermostable esterase, EstE1, conserved catalytic triad made up of Ser, Asp, and His, resembled greater evolutionary affinity with carboxylesterase from S. roseus45. Bourne et al. reported the pentapeptide consensus sequence (Gly-X1-Ser-X2-Gly) around the catalytic serine residue46. The catalytic machinery of esterase depended on the integrity of the catalytic dyad or triad. Richter et al. computationally designed esterase with a Cys-His dyad and an oxygen anion hole in the active site47. They found that, besides the catalytic triad residues (Ser-His-Glu), oxyanion hole stabilized the negative charge on the carbonyl oxygen of the tetrahedral intermediate. In most cases, serine carried the nucleophilic hydroxyl group, histidine acted as a general base and phenylalanine provided acidic moiety47. Previous studies reported that carboxylesterase has the ability to metabolize a broad range of insecticides14,44. In recent study, we found that carboxylesterase from S. roseus possessed relatively broad variety of pesticides degrading ability.
Materials and methods
Genomic context of pesticide degradation
Considering the ability of esterase mediated degradation of pesticides from this bacterial strain, the genes related to the degradation was checked in the genome of S. roseus strain RF1H. For this, the whole genome sequencing was carried-out on the Illumina MiSeq platform from ICRISAT Patancheru, Hyderabad, India. The assembled genome was visualized using the CGView server (https://cgview.ca/). Further, circular map of selected genomes of S. roseus strains (MOSEL-ME25, CCM 3516, DSM 5351, BU-1 and W12; accessed on May 17, 2022), with comparative alignment against strain RF1H were generated using CGView server48. The predict genes, protein functions and subsystem categories were annotated using Rapid Annotations using Subsystems Technology (RAST), and The SEED viewer49,50.
Previously, the 16S rRNA gene sequence of S. roseus strain RF1H was submitted to NCBI nucleotide database with the Accession number MN080417. Bacterial culture was deposited to National Center for Microbial Resource, Pune, India (Accession number MCC 4829) and Microbial Type Culture Collection and Gene Bank, Chandigarh, India (Accession number MTCC13151).
Chemicals and the preparation of enzyme
All the chemicals and media components were purchased from HiMedia Laboratory Ltd., Mumbai, India. In addition, five groups of pesticides viz., Hexaconazole, Mancozeb, Pretilachlor, Organophosphate and λ-cyhalothrin were received from Crop Research and Seed Multiplication Farm (CRSMF) of Burdwan University.
Crude esterase was procured from previously isolated bacterium Salinicoccus roseus strain RF1H in optimized media38. Crude esterase solution was partially purified by addition of ammonium sulfate [(NH4)2SO4] precipitation (up to 70%) followed by dialysis at 4 °C for 24 h. Protein content of the partially purified enzyme was determined spectrophotometrically by Bradford assay, taking bovine serum albumin (BSA) as a reference standard51.
Estimation of enzyme activity
Esterase activity was determined using p-nitrophenyl acetate (p-NPA) as the substrate, following Winkler and Stuckmann method with modification52. Briefly, the assay mixture was comprised of crude esterase (0.5 mL), p-NPA (0.5 mL) as a reaction substrate (10mM dissolved in Ethanol), 50mM phosphate buffer pH 7.0 (3.5 mL) and incubated at 37 °C for 15 min. The control sample (blank) was comprised only of p-NPA and buffer solution.
A spectrophotometer [Labtronics, LT 39, India] was used to record the absorbance at the wavelength of 405 nm. The absorbance of blank samples (excluding enzyme) was recorded using similar way. Enzyme activity was expressed as international units (IU), where 1 IU represented the amount of enzyme that released 1 µmol of product per minute under defined assay conditions.
Beads fabrication
Gel beads were prepared following Ahmed et al. with slight modification53. For this, partially purified esterase was immobilized by mixing with an equal volume (1:1) of sodium alginate solution [2.5% (w/v)] by continuous stirring. The homogenous mixture was extruded dropwise through a hypodermic syringe into 50mL calcium chloride solution 1% (w/v) and hardened for 1 h. The spherical beads were harvested by filtration, rinsed with deionized water, and left for air drying overnight. The activity of immobilized esterase was estimated as described above, except that 0.5 g of beads was taken for assay. One unit of esterase activity was defined as the amount of esterase that yielded 1 µmol of p-nitrophenyl per mL min− 1 at 37 °C temperature.
Characterization of beads
The surface morphology of the alginate bead before and after esterase immobilization was studied by using a scanning electron microscope (SEM) (S-530, Hitachi, Japan). The beads were mounted in SEM stub and coated with gold by sputtering and observed at an electron high tension (EHT) of 5 kV. The Fourier-transform infrared (FTIR) spectra were measured with an FTIR spectrophotometer (Cary-630, Agilent, USA) with a scanning range of 400–4500 cm− 1. Crystalline properties of the beads were examined by X-ray diffractometer (XRD) (X-PERT PRO, Malvern Panalytical, UK) with a 2θ angle ranging from 10 to 90° and a wavelength of 0.154 nm, at 45 kV and 40 mA. Thermal stability was measured by means of thermogravimetric analysis (TGA) (Perkin Elmer, USA). The temperature was ramped from 20 to 392 °C at a heating rate of 10 °C/min under a nitrogen atmosphere.
Pesticides degradation
100 mg/L of different tested pesticides like Hexaconazole, Mancozeb, Pretilachlor, Organophosphate and λ-cyhalothrin were reacted with equivalent amount (0.420 U/mL) of free esterase and immobilized esterase in presence of tributyrin (1% v/v), at pH 5.0, incubation temperature of 37 °C and shaking speed of 150 rpm for four days. Residual pesticides were estimated by taking absorbance at 410 nm by Digital Spectrophotometer [Labtronics, LT 39, India]. Degradation rate was calculated by the following formula:
Where, ‘mm’ was the absorbance of pesticide solution at initial stage and ‘mt’ was the absorption of pesticide solution after four days.
Chromatographic analysis
Residuals of the pesticide (Hexaconazole) were extracted from the supernatant using equal volume of ethyl acetate and acetone (1:1) (analytical grade). The extraction procedure was repeated three times. The solvent was removed using rotary evaporator [BioBase Re 100-Pro, China] and the final residue was dissolved in 2 mL methanol. The samples were analyzed using liquid chromatography-mass spectrometry (LC-MS) [Agilent 6545 Q-TOF LC/MS, Agilent Technologies, USA] to find out the degradation intermediates of pesticides. The quadrupole Time of Flight LC/MS system was equipped with electrospray ionization (ESI) and operated in the positive ion mode (175.0 V). The non-polar octadecylsilane phase (C18) was used as the stationary phase and the mobile phase consisted of solvents in 80:20 acetonitrile/water (Milli-Q purified water) at a flow rate of 0.25 mL/min and the column temperature was 60 °C. A sample volume of 3 µL was injected onto the column [ZORBAX Eclipse Plus C18, 2.1 × 150 mm, 1.8 μm]. The Q-TOF detector scan range was from 50 to 1000 m/z. The analysis time for LC-MS was 15 min.
Root growth and root tip preparation
Onion bulbs (Allium cepa L.) (2n = 16) were placed in test tubes and rooted in distilled water for 24 h. On the emergence of new roots, three onion bulbs were selected and suspended on the beakers. Three different experimental set up were cultivated in the beaker, containing control (distilled water), Hexaconazole solution (100 mg/L) and pesticide degraded solution, and kept for 72 h.
For the purpose of cytotoxicity test, root tips of 1–3 cm long were collected and stained with Aceto-orcein, following standard protocol36. The comparison of toxic effects was studied by calculating the mitotic index (MI) using the following formula. Chromosomal aberrations in different stages of mitosis were viewed under the light microscope (Leica, DM750, Germany).
Kinetic behavior of free and immobilized esterase
The catalytic parameters like maximum velocity (Vmax) and Michaëlis-Menten constant (KM), and kcat of free and immobilized esterase were calculated from the Lineweaver and Burk plot at standard assay conditions54.Various concentrations of substrate para-Nitrophenyl acetate (p-NPA) ranging from 1mM to 10mM were used. Lineweaver-Burk equation has been given as follows-
Where, (1/V) and (1/S) were the reciprocal velocity and substrate concentration respectively. Michaëlis-Menten kinetics was obtained from the slope and intercept of a plot of 1/[V] versus 1/[S]. The catalytic constant (kcat) was determined using the following equation:
Where, [E] was the total amount of enzyme in the reaction mixture.
Molecular modelling, docking and dynamics simulation study
The three-dimensional structure of the selected protein, esterase from Salinicoccus roseus was not yet available in RCSB PDB database [https://www.rcsb.org/], thus homology modelling technique was employed in the retrieved sequence [ID A0A0C2HD44] from UniProt database [https://www.uniprot.org/] using SWISS- MODEL Workspace [https://swissmodel.expasy.org/interactive]. The crystal structure with highest sequence similarity was selected as template [8FTP] for esterase. Furthermore, the stereochemical quality of the refined model was assessed by PROCHECK [https://servicesn.mbi.ucla.edu/PROCHECK/], ERRAT [https://servicesn.mbi.ucla.edu/ERRAT/] and VERIFY 3D [https://servicesn.mbi.ucla.edu/viewer/?job=9678andp=verify3d]. Finally, Z-score of the protein was calculated by PROVE server [https://saves.mbi.ucla.edu/]. The optimized model served as the target enzyme for further analysis. 3D structure of pesticides viz., Hexaconazole [CID 66461], Mancozeb [CID 300605], Pretilachlor [CID 91644], Organophosphate [CID 1982] and λ-cyhalothrin [CID 6440557] were retrieved from PubChem database [https://pubchem.ncbi.nlm.nih.gov/] in spatial data file (SDF) format. Discovery Studio software (version 17.2.0.16349, 2017 R2) was used for further conversion into protein data bank (PDB) format before docking. The active site of the modeled protein was identified by Computed Atlas of Surface Topography of proteins (CASTp) 3.0 tool. A computational ligand-target docking approach was carried out in AutoDock 4.2.6 program. Only polar hydrogen atoms were added by Gasteiger method, and Kollaman charges were applied to the target molecule. Docking simulation was performed by Lamarckian genetic algorithm (LGA) with 10 independent docking runs on Cygwin with maximum energy evaluations of 2,500,00055. The dimensions of the grid box [60 × 40 × 50] were pointing in the x, y and z directions, respectively, with a grid point spacing of 0.0375 nm in AutoGrid 4. After docking, the lowest binding energy (Kcal/mol) was selected for subsequent analysis. In order to evaluate the robustness of the systems under investigation, the Root Mean Square Deviation (RMSD) was computed as a metric for quantifying the extent of deviation in atomic positions from their respective initial coordinates throughout the entire simulation period. During simulation, the RMSD was calculated of the coordinate of before simulation with after simulation-complex file.MD simulation of modeled esterase protein was performed by using GROMACS 2021.1 software package with Gromos 54a7 force field and the flexible SPC water model56,57. The initial structure was immersed in a periodic water box of cubic shape (0.5 nm thick). Electrostatic energy was calculated using the particle mesh Ewald method, which permits the use of Ewald summation at a computational cost comparable to that of a simple truncation method of 1 nm or less58. We retained the cutoff distance as 1.0 nm for the calculation of the coulomb and van der Waal’s interaction, respectively. After energy minimization using the steepest descent for 1000 steps, the system was subjected to equilibration at 300 k and normal pressure for 100 ps under the conditions of position restrainted for heavy atoms. We subsequently applied LINCS constrainted for all bonds, keeping the whole protein molecule fixed and allowing only the water molecule to move to equilibrate with respect to the protein structure59. The system was coupled to the external bath by the Berendsen pressure and temperature coupling59.
At initial, 1ns of the five complexes were analysed to validate the stability of the receptor, based on structural coordinate deviation.The final MD calculations were performed for 100 ns under the same conditions except that the position restrainted were removed. The results were analyzed using the standard software provided by the GROMACS package. An average structure was further refined using the steepest descent energy minimization. Simulated complexes were utilized to produce bond interaction using the Discovery Studio Visualization system.
Statistical analysis
A two-way ANOVA, followed by Tukey HSD test (p < 0.05) was performed to determine the statistical significance of the effects of immobilized esterase and free esterase on pesticide degradation, using R software version 4.2.060. The level of significance for the MI was evaluated by One-way ANOVA followed by Post Hoc test60. All tests were conducted in triplicate.
Conclusions
In the current study, entrapment of esterase in sodium alginate beads has proven a successful, simple, and inexpensive method of enzyme immobilization. Immobilized esterase has been employed for the biodegradation of pesticides because it conferred the highest catalytic activity for a long time. The complexes obtained from molecular docking provided a better understanding of the degradation processes performed by esterase. The results reinforced the importance of molecular docking studies for enzyme activity comprehension. The carboxylesterase gene was not located in the genomic island thus suggesting the gene was acquired through evolution and was associated with hydrocarbon degradation. Therefore, experimentally determined results and subsequent in silico evidences provided atomistic knowledge of their binding mechanism, and opens up possibility to design biopesticides. Toxicity test results indicated that pesticide degraded solution is non-toxic and helps in A. cepa cell growth. This suggests that esterase mediated biodegradation could be an alternative solution of pesticide pollution. These information can allow in the future implementation of new strategies for pesticide management. In the field of pesticide bioremediation, recombinant DNA technology can be an optional tool to develop Genetically engineered organisms (GEM) tailored with pesticide degrading genes through specific enzyme. Use of effective methods like transcriptional modification of important metabolic gene and regulator, nanoformulation of pesticides, and novel carriers can be a vehicle of green technology. Hence, immobilized esterase on alginate beads can be potential for reducing the pesticide damage. This research work scintillates the practical application assisting bioelimination.
Data availability
All the generated data are available with the article.
Abbreviations
- ANOVA:
-
analysis of variance
- BSA:
-
bovine serum albumin
- FTIR:
-
fourier-transform infrared spectroscopy
- GEM:
-
genetically engineered organisms
- IU:
-
international units
- K M :
-
michaelis constant
- k cat :
-
catalytic constant
- LC-MS:
-
liquid chromatography–mass spectrometry
- MD:
-
molecular dynamics
- MI:
-
mitotic index
- (NH4)2SO4 :
-
Ammonium sulfate
- OC:
-
organochlorines
- OP:
-
organophosphates
- p-NPA:
-
p-nitrophenyl acetate
- POP:
-
persistent organic pollutants
- RMSD:
-
root mean square deviation
- SEM:
-
scanning electron microscopy
- TGA:
-
thermogravimetric analysis
- V max :
-
maximal velocity
- XRD:
-
X-ray diffractometer
References
Vashistha, V. K. et al. Stereoselective analysis of chiral pesticides: a review. Environ. Monit. Assess.196, 1–9. https://doi.org/10.1007/s10661-024-12310-0 (2024).
Özkara, A., Akyıl, D. & Konuk, M. Pesticides, environmental pollution, and health. Environmental health risk-hazardous factors to living species. IntechOpen (2016). https://doi.org/10.5772/63094
World Health Organization. Guidelines for establishing a poison centre. Geneva. (2020). https://www.who.int/publications/i/item/9789240009523
Chandra, R., Sharpanabharathi, N., Prusty, B. A., Azeez, P. A. & Kurakalva, R. M. Organochlorine pesticide residues in plants and their possible ecotoxicological and agri food impacts. Sci. Rep.11, 17841. https://doi.org/10.1038/s41598-021-97286-4 (2021).
Bawa, P. et al. Persistent organic pollutants residues in human breast milk from Bathinda and Ludhiana districts of Punjab, India. Arch. Environ. Contam. Toxicol.75, 512–520. https://doi.org/10.1007/s00244-018-0512-3 (2018).
SalehI.A., Zouari, N. & Al-Ghouti, M. A. Removal of pesticides from water and wastewater: Chemical, physical and biological treatment approaches. Environ. Technol. Innov.101026https://doi.org/10.1016/j.eti.2020.101026 (2020). (2020).
Jayaraj, R., Megha, P. & Sreedev, P. Organochlorine pesticides, their toxic effects on living organisms and their fate in the environment. Interdiscip Toxicol.9, 90–100. https://doi.org/10.1515/intox-2016-0012 (2016).
Bielská, L., Hale, S. E. & Škulcová, L. A review on the stereospecific fate and effects of chiral conazole fungicides. Sci. Total Environ.750, 141600. https://doi.org/10.1016/j.scitotenv.2020.141600 (2020).
Černohlávková, J., Jarkovský, J. & Hofman, J. Effects of fungicides mancozeb and dinocap on carbon and nitrogen mineralization in soils. Ecotoxicol. Environ. Saf.72, 80–85. https://doi.org/10.1016/j.ecoenv.2008.07.001 (2009).
Shilpakar, O., Karki, B. & Rajbhandari, B. Pretilachlor poisoning: a rare case of a herbicide masquerading as organophosphate toxicity. Clin. Case Rep.8, 3507–3509. https://doi.org/10.1002/ccr3.3473 (2020).
Fu, H. et al. Advances in organophosphorus pesticides pollution: current status and challenges in ecotoxicological, sustainable agriculture, and degradation strategies. J. Hazard. Mater.424, 127494. https://doi.org/10.1016/j.jhazmat.2021.127494 (2022).
Wendeborn, S., Godineau, E., Mondière, R., Smejkal, T. & Smits, H. Chirality in Agrochemicals in Comprehensive Chirality (ed. Carreira, E.M., Yamamoto, H.) 120–166. https://doi.org/10.1016/B978-0-08-095167-6.00102-6 (Elsevier, 2012).
Zheng, J. Y. et al. A stereoselective esterase from Bacillus megaterium: purification, gene cloning, expression and catalytic properties. Protein Expr Purif.136, 66–72. https://doi.org/10.1016/j.pep.2015.10.001 (2017).
Zuo, Z. et al. Engineering Pseudomonas putida KT2440 for simultaneous degradation of organophosphates and pyrethroids and its application in bioremediation of soil. Biodegradation26, 223–233. https://doi.org/10.1007/s10532-015-9729-2 (2015).
Zdarta, J., Meyer, A. S., Jesionowski, T. & Pinelo, M. Multi-faceted strategy based on enzyme immobilization with reactant adsorption and membrane technology for biocatalytic removal of pollutants: a critical review. Biotechnol. Adv.37, 107401. https://doi.org/10.1016/j.biotechadv.2019.05.007 (2019).
Araújo, P. D. S., Belini, G. B., Mambrini, G. P., Yamaji, F. M. & Waldman, W. R. Thermal degradation of calcium and sodium alginate: a greener synthesis towards calcium oxide micro/nanoparticles. Int. J. Biol. Macromol.140, 749–760. https://doi.org/10.1016/j.ijbiomac.2019.08.103 (2019).
Guzik, U., Hupert-Kocurek, K., Marchlewicz, A. & Wojcieszynska, D. Enhancement of biodegradation potential of catechol 1, 2-dioxygenase through its immobilization in calcium alginate gel. Electron. J. Biotechnol.17, 83–88. https://doi.org/10.1016/j.ejbt.2014.02.001 (2014).
Hou, Y. et al. Applying molecular docking to pesticides. Pest Manag Sci.79, 4140–4152. https://doi.org/10.1002/ps.7700 (2023).
Adrio, J. L. & Demain, A. L. Recombinant organisms for production of industrial products. Bioeng. Bugs.1, 116–131. https://doi.org/10.4161/bbug.1.2.10484 (2010).
Richins, R. D., Mulchandani, A. & Chen, W. Expression, immobilization, and enzymatic characterization of cellulose-binding domain‐organophosphorus hydrolase fusion enzymes. Biotechnol. Bioeng.69, 591–596. https://doi.org/10.1002/1097-0290%2820000920%2969:6%3C591::AID-BIT2%3E3.0.CO;2-X (2000).
Xiao, Y., Lu, Q., Yi, X., Zhong, G. & Liu, J. Synergistic degradation of pyrethroids by the quorum sensing-regulated carboxylesterase of Bacillus subtilis BSF01. Front.Bioeng. Biotechnol.8, 889. https://doi.org/10.3389/fbioe.2020.00889 (2020).
Bhatt, P., Zhou, X., Huang, Y., Zhang, W. & Chen, S. Characterization of the role of esterases in the biodegradation of organophosphate, carbamate, and pyrethroid pesticides. J. Hazard. Mater.411, 125026. https://doi.org/10.1016/j.jhazmat.2020.125026( (2021).
Lemes, A. C., Silvério, S. C., Rodrigues, S. & Rodrigues, L. R. Integrated strategy for purification of esterase from Aureobasidium pullulans. Sep. Purif. Technol.209, 409–418. https://doi.org/10.1016/j.seppur.2018.07.062 (2019).
Xin, L. & Hui-Ying, Y. Purification and characterization of an extracellular esterase with organic solvent tolerance from a halotolerant isolate, Salimicrobium sp. LY19. BMC Biotechnol.13, 1–8. https://doi.org/10.1186/1472-6750-13-108 (2013).
He, F., Zhang, S. & Liu, X. Immobilization of feruloyl esterases on magnetic nanoparticles and its potential in production of ferulic acid. J. Biosci. Bioeng.120, 330–334. https://doi.org/10.1016/j.jbiosc.2015.01.006 (2015).
Gul, S., Khan, S. B., Rehman, I. U., Khan, M. A. & Khan M.I. A comprehensive review of magnetic nanomaterials modern day theranostics. Front. Mater.6, 179. https://doi.org/10.3389/fmats.2019.00179 (2019).
Tercan, Ç. & Sürmeli, Y. Şanlı-Mohamed, G.Thermoalkalophilic recombinant esterase entrapment in chitosan/calcium/alginate‐blended beads and its characterization. J. Chem. Technol. Biotechnol.96, 2257–2264. https://doi.org/10.1002/jctb.6750 (2021).
Raghu, S. & Pennathur, G. Enhancing the stability of a carboxylesterase by entrapment in chitosan coated alginate beads. Turk. J. Biol.42, 307–318. https://doi.org/10.3906/biy-1805-28 (2018).
Gülay, S. & Şanlı-Mohamed, G. Immobilization of thermoalkalophilic recombinant esterase enzyme by entrapment in silicate coated Ca-alginate beads and its hydrolytic properties. Int. J. Biol. Macromol.50, 545–551. https://doi.org/10.1016/j.ijbiomac.2012.01.017 (2012).
Soares, J. D., Santos, J. E., Chierice, G. O. & Cavalheiro, E. T. Thermal behavior of alginic acid and its sodium salt. Eclét Quím. 29, 57–64. https://doi.org/10.1590/S0100-46702004000200009 (2004).
Wasupalli, G. K. & Verma, D. Molecular interactions in self-assembled nano-structures of chitosan-sodium alginate based polyelectrolyte complexes. Int. J. Biol. Macromol.114, 10–17. https://doi.org/10.1016/j.ijbiomac.2018.03.075 (2018).
Daemi, H. & Barikani, M. Synthesis and characterization of calcium alginate nanoparticles, sodium homopolymannuronate salt and its calcium nanoparticles. Sci. Iran.19, 2023–2028. https://doi.org/10.1016/j.scient.2012.10.005 (2012).
Mali, H., Shah, C., Patel, D. H., Trivedi, U. & Subramanian, R. B. Bio-catalytic system of metallohydrolases for remediation of neurotoxin organophosphates and applications with a future vision. J. Inorg. Biochem.https://doi.org/10.1016/j.jinorgbio.2022.111771( (2022).
Yu, S. J. Detoxification mechanisms in insects in Encyclopedia of Entomology (ed Capinera, J. L.) 1187–1201. https://doi.org/10.1007/978-1-4020-6359-6_891 (Springer, (2008).
Singh, B., Kaur, J. & Singh, K. Biodegradation of malathion by Brevibacillus sp. strain KB2 and Bacillus cereus strain PU. World J. Microbiol. Biotechnol.28, 1133–1141. https://doi.org/10.1007/s11274-011-0916-y (2012).
Mercado, S. A. & Caleño, J. D. Cytotoxic evaluation of glyphosate, using Allium cepa L. as bioindicator. Sci. Total Environ. 700–134452. https://doi.org/10.1016/j.scitotenv.2019.134452 (2020).
Mesi, A. & Kopliku, D. Cytotoxic and genotoxic potency screening of two pesticides on Allium cepa L. Proc. Technol. 8,19–26. (2013). https://doi.org/10.1016/j.protcy.2013.11.005
Dutta, B., Nigam, V. K., Panja, A. S., Shrivastava, S. & Bandopadhyay, R. Statistical optimisation of esterase from Salinicoccus roseus strain RF1H and its potential application in synthetic dye decolorisation. Biocatal. Biotransfor. 41, 67–80. https://doi.org/10.1080/10242422.2021.2010718 (2021).
Rodrigues, R. C., Ortiz, C., Berenguer-Murcia, Á., Torres, R. & Fernández-Lafuente, R. Modifying enzyme activity and selectivity by immobilization. Chem. Soc. Rev.42, 6290–6307. https://doi.org/10.1039/c2cs35231a (2013).
Aslam, S. et al. Fabrication and catalytic characterization of laccase-loaded calcium-alginate beads for enhanced degradation of dye-contaminated aqueous solutions. Catal. Lett.152, 1729–1741. https://doi.org/10.1007/s10562-021-03765-8 (2022).
Wei, T. et al. Characterization of a novel thermophilic pyrethroid-hydrolyzing carboxylesterase from Sulfolobus tokodaii into a new family. J. Mol. Catal. B Enzym. 97, 225–232. https://doi.org/10.1016/j.molcatb.2013.07.022 (2013).
Karakuş, E., Özler, A. & Pekyardimci, Ş. Noncovalent immobilization of pectinesterase (Prunus armeniaca L.) onto bentonite. Artif. Cell. Blood Sub. 36, 535–550. https://doi.org/10.1080/10731190802554455 (2008).
Page, M. I. & 332 The energetics of intramolecular reactions and enzyme catalysis. Phil. Trans. R. Soc. Lond. B. (1991). 149 – 56. https://doi.org/10.1098/rstb.1991.0043.
Wang, B. Z. et al. Cloning of a novel pyrethroid-hydrolyzing carboxylesterase gene from Sphingobium sp. strain JZ-1 and characterization of the gene product. Appl. Environ. Microbiol.75, 5496–5500. https://doi.org/10.1128/AEM.01298-09 (2009).
Byun, J. S. et al. Crystal structure of hyperthermophilic esterase EstE1 and the relationship between its dimerization and thermostability properties. BMC Struct. Biol.7, 47. https://doi.org/10.1186/1472-6807-7-47 (2007).
Bourne, P. C., Isupov, M. N. & Littlechild, J. A. The atomic-resolution structure of a novel bacterial esterase. Structure. 8https://doi.org/10.1016/S0969-2126(00)00090-3 (2000). 143 – 51.
Richter, F. et al. Computational design of catalytic dyads and oxyanion holes for ester hydrolysis. J. Am. Chem. Soc.134, f16197–f16206. https://doi.org/10.1021/ja3037367 (2012).
Grant, J. R. & Stothard, P. The CGView server: a comparative genomics tool for circular genomes. Nucleic Acids Res.36, W181–184. https://doi.org/10.1093/nar/gkn179 (2008).
Aziz, R. K. et al. The RAST server: rapid annotations using subsystems technology. BMC Genom.9, 1–5. https://doi.org/10.1186/1471-2164-9-75 (2008).
Overbeek, R. et al. The SEED and the Rapid Annotation of microbial genomes using Subsystems Technology (RAST). Nucleic Acids Res.42, D206–214. https://doi.org/10.1093/nar/gkt1226 (2014).
Bradford, M. M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem.72, 248–254. https://doi.org/10.1006/abio.1976.9999 (1976).
Winkler, U. K., Stuckmann, M. A. & Glycogen Hyaluronate, and some other polysaccharides greatly enhance the formation of exolipase by Serratia marcescens. J. Bacteriol.138, 663–670. https://doi.org/10.1128/jb.138.3.663-670.1979 (1979).
Ahmed, N. E., El Shamy, A. R. & Awad, H. M. Optimization and immobilization of amylase produced by aspergillus terreus using pomegranate peel waste. Bull. Natl. Res. Cent.44 (1-2). https://doi.org/10.1186/s42269-020-00363-3 (2020). (2020).
Lineweaver, H. & Burk, D. The determination of enzyme dissociation constants. J. Am. Chem. Soc.56, 658–666. https://doi.org/10.1021/JA01318A036 (1934).
Fuhrmann, J., Rurainski, A., Lenhof, H. P. & Neumann, D. A new lamarckian genetic algorithm for flexible ligand-receptor docking. J. Comput. Chem.31, 1911. https://doi.org/10.1002/jcc.21478 (2010).
Hess, B., Kutzner, C., Spoel, D. V. D. & Lindahl, E. GROMACS 4: algorithms for highly efficient, load-balanced, and scalable molecular simulation. J. Chem. Theory Comput.4, 435–447. https://doi.org/10.1021/ct700301q (2008).
Oostenbrink, C., Villa, A., Mark, A. E. & van Gunsteren, W. F. A biomolecular force field based on the free enthalpy of hydration and solvation: the GROMOS force-field parameter sets 53A5 and 53A6. J. Comput. Chem.25, 1656–1676. https://doi.org/10.1002/jcc.20090 (2004).
Wells, B. A. & Chaffee, A. L. Ewald summation for molecular simulations. J. Chem. Theory Comput.11, 3684–3695. https://doi.org/10.1021/acs.jctc.5b00093 (2015).
Perez-Sanchez, G., Galvão, T. L., Tedim, J. & Gomes, J. R. A molecular dynamics framework to explore the structure and dynamics of layered double hydroxides. Appl. Clay Sci.163, 164–177. https://doi.org/10.1016/j.clay.2018.06.037 (2018).
R Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria.URL (2021). http://www.R-project.org/
Acknowledgements
Financial assistance to BD in the form of Senior Research Fellow (SRF) received from Department of Science and Technology (DST), New Delhi, India for PURSE Phase II program is gratefully acknowledged. We thank BIT, Mesra, Ranchi, Jharkhand and NIPER, Kolkata, West Bengal for their support during the present study. We extend our thanks to Mr. Rajdeep Shaw, Research Scholar, University of Burdwan for his help during the mitotic study.
Funding
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Author information
Authors and Affiliations
Contributions
RB: Conceptualization, supervision. BD: Execution, analysis, draft manuscript writing ASP: Software, Ligand preparation and MD simulation. VKN: Review, editing SN: Chromatography work and review, VR: Chromatography. All authors reveiwed and agreed to manuscript publication.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Consent to publish
All authors agreed to manuscript publication.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Dutta, B., Panja, A.S., Nigam, V.K. et al. Computational and biochemical characterization of the immobilized esterase of Salinicoccus roseus for pesticide degradation. Sci Rep 14, 30661 (2024). https://doi.org/10.1038/s41598-024-73165-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-024-73165-6
